|
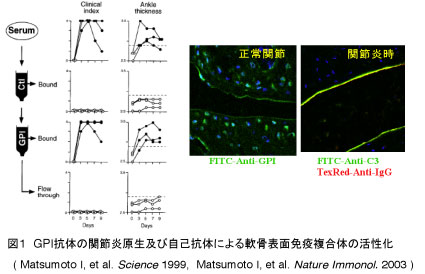
怴婯娭愡墛儌僨儖儅僂僗傪梡偄偨尋媶 娭愡儕僂儅僠乮RA乯偵懳偡傞帯椕愴棯偼丄庮釃夡巰場巕乮TNF兛乯丄僀儞僞乕儘僀僉儞-6 (IL-6)側偳偺墛徢惈僒僀僩僇僀儞偺嶌梡傪梷惂偡傞暘巕昗揑帯椕栻偺奐敪偵傛傝戝偒偔曄慗偟傑偟偨丅偟偐偟側偑傜丄RA徢椺偺栺3妱偼TNF慾奞椕朄偵掞峈惈偱丄怴婯峈儕僂儅僠栻偺奐敪偼昁帄偱偡丅変乆偼丄僸僩偺RA昦懺偵傛傝嬤偄娭愡墛儌僨儖儅僂僗傪梡偄偰丄峏側傞怴婯帯椕朄傪栚巜偟偰尋媶傪峴偭偰偄傑偡丅 亂Glucose-6-phosphate isomerase (GPI)桿摫娭愡墛儌僨儖偲偼亃 GPI偼儐價僉僞僗偵懚嵼偡傞夝摐宯峺慺偺堦偮偱偁傞偙偲偑抦傜傟偰偄傑偡偑丄帺慠敪徢娭愡墛儌僨儖偱偁傞K/BxN儅僂僗偺帺屓峈尨偲偟偰摨掕偝傟丄GPI偺娭愡墛尨惈偑帵偝傟傑偟偨乮恾侾乯丅
亂GPI桿摫娭愡墛儌僨儖偵偍偗傞IL-6/IL-17偺栶妱亃 RA姵幰娭愡妸枌偵偼丄CD4梲惈T嵶朎偑懡偔怹弫偟丄堦晹偺T嵶朎偑僋儘乕僫儖偵憹怋偡傞偙偲偑抦傜傟偰偄傑偡丅GPI桿摫娭愡墛偺敪徢偵傕CD4梲惈T嵶朎偑昁恵偱偁傞偙偲偼丄2004擭Schubert傜偵傛偭偰柧傜偐偵偝傟傑偟偨丅傢傟傢傟偼偙偺GPI桿摫娭愡墛儅僂僗偵峈IL-17拞榓峈懱傪搳梌偡傞偙偲偵傛傝娭愡墛傪桳堄偵梷惂偡傞偙偲偑偱偒傞偙偲偐傜丄娭愡墛偺敪徢偵偼Th17嵶朎偑婑梌偟偰偄傞偙偲傪曬崘偟傑偟偨丅傑偨丄峈IL-6庴梕懱峈懱搳梌偼Th17嵶朎偺暘壔慾奞丄帺屓峈懱偺嶻惗梷惂丄峈尨摿堎揑T嵶朎偺憹怋梷惂傪傕偨傜偡偙偲偵傛傝帺屓柶塽惈娭愡墛偑惂屼偝傟丄尰嵼偺IL-6梷惂帯椕偺儊僇僯僘儉偺堦抂傪柧傜偐偵偟傑偟偨乮恾3乯丅
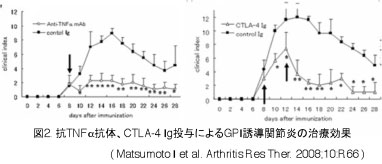
亂GPI儁僾僠僪桿摫娭愡墛儌僨儖偺妋棫亃 GPI偼558傾儈僲巁偐傜峔惉偝傟偰偄傞僞儞僷僋幙偱偡丅巹偨偪偼GPI桿摫娭愡墛偺T嵶朎僄僺僩乕僾傪摨掕偟丄娭愡墛桿摫偵廳梫側僄僺僩乕僾偑丄GPI325-339偱偁傞偙偲傪柧傜偐偵偟傑偟偨丅GPI325-339儁僾僠僪傪柶塽偟丄day0偲day2偵昐擔奝撆慺乮PT乯傪暊峯撪搳梌偡傞偙偲偱丄GPI桿摫娭愡墛儌僨儖偲摨條偵柶塽屻1廡娫掱搙偐傜憗婜偵娭愡墛傪庝婲偱偒傑偡丅偙傟偼悽奅偱弶傔偰偺儁僾僠僪傪梡偄偨娭愡墛桿摫儌僨儖偱偡丅
忋婰偵帵偟偨傛偆偵GPI桿摫娭愡墛偵偼敪徢帪婜偵傛傜偢CD4梲惈T嵶朎偺栶妱偑廳梫偱偁傞偙偲偑偡偱偵帵偝傟偰偄傞偙偲丄GPI偺T嵶朎僪儈僫儞僩僄僺僩乕僾偺摨掕偐傜丄峈尨摿堎揑CD4梲惈T嵶朎偵徟揰傪偍偄偨尋媶傕恑傔偰偍傝丄娭愡墛偵娭梌偟偰偄傞偲峫偊傜傟傞怴婯昦場揑暘巕偺扵嶕偲偦偺儊僇僯僘儉偵偮偄偰夝愅傪恑傔偰偄傑偡丅 怴婯娭愡墛梷惂暘巕(TIARP / STEAP4)偵娭偡傞尋媶
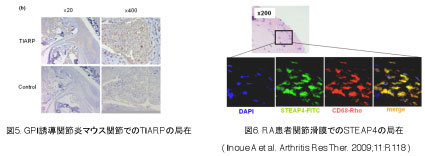
亂GPI桿摫娭愡墛儌僨儖偵偍偗傞怴婯昦場揑場巕偺夝柧亃 RA偵偍偗傞TNF兛偺廳梫惈偼丄悢乆偺椪彴僨乕僞偐傜柧傜偐偵偝傟偰偄傑偡丅変乆偼丄RA偍傛傃GPI桿摫娭愡墛偱昦懺丒昦場偵嫮偔娭梌偟偰偄傞TNF兛偵徟揰傪偍偒丄娭愡墛敪徢捈屻偺儅僂僗銪憻偵偍偗傞TNF娭楢堚揱巕偺敪尰曄摦傪Gene Chip偱夝愅偟傑偟偨丅偦偺寢壥丄TNF娭楢暘巕偺偆偪丄TNF兛-induced adipose-related protein (TIARP)暘巕偑懠偺TNF娭楢場巕偲斾傋偰娭愡墛儅僂僗銪憻偱柧傜偐偵嫮敪尰偟偰偄傞偙偲偑暘偐傝傑偟偨丅TIARP偼丄帀朾嵶朎偺暘壔偵娭梌偡傞暘巕偲偟偰弶傔偵摨掕偝傟丄TNF兛丄IL-6傗IL-1偵傛傝桿摫偝傟傞偲偺曬崘偑側偝傟偰偄傑偡丅TIARP偺僸僩ortholog偲偟偰偼STEAP4偑摨掕偝傟偰偄傑偡偑丄儅僂僗丄僸僩偲傕偵幚嵺偺惗暔妛揑側婡擻丄傑偨娭愡墛傪娷傓帺屓柶塽惈幘姵偲偺娭梌偺曬崘偼偁傝傑偣傫丅変乆偺夝愅偵傛傝TIARP偼娭愡墛儅僂僗銪憻偍傛傃娭愡偵嬊嵼偟丄摿偵CD11b梲惈偺儅僋儘僼傽乕僕偍傛傃憹怋偟偨妸枌嵶朎偵嫮偔敪尰偡傞偙偲傪尒偄偩偟傑偟偨乮恾1A乯丅峏偵変乆偼僸僩RA姵幰偺妸枌慻怐偵偍偄偰傕TIARP偺僸僩ortholog偱偁傞STEAP4敪尰傪妋擣偟偰偄傑偡乮恾1B乯丅
恾1. GPI桿摫娭愡墛偍傛傃RA姵幰娭愡妸枌偵偍偗傞TIARP/STEAP4偺嬊嵼
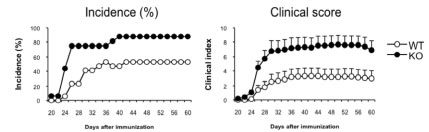
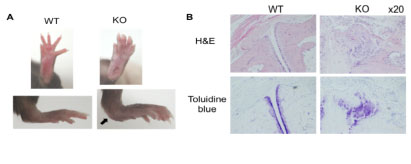
亂TIARP偺帺屓柶塽惈娭愡墛偵偍偗傞昦場揑堄媊亃 娭愡墛儅僂僗銪憻偍傛傃娭愡偵嫮敪尰偡傞TIARP偺娭愡墛偵偍偗傞婡擻傪柧傜偐偵偡傞偨傔偵傢傟傢傟偼丄撈帺偵TIARP寚懝儅僂僗傪嶌惢偟丄娭愡墛偵偍偗傞TIARP暘巕偺堄媊傪傛傝徻嵶偵専摙偟傑偟偨丅 TIARP寚懝儅僂僗 (C57BL/6宯摑) 偵僐儔乕僎儞桿摫娭愡墛(CIA)偺桿摫傪帋傒傞偲丄TIARP寚懝儅僂僗偱偼CIA偺敪徢棪丄廳徢搙偲傕偵栰惗宆儅僂僗偲斾妑偟偰桳堄偵埆壔偡傞偙偲偑傢偐傝傑偟偨(恾2A)丅娭愡偺慻怐妛揑僗僐傾(墛徢搙丄崪傃傜傫搙)傕挊柧偵崅偔丄墛徢晹埵偵偼懡悢偺岲拞媴丄M兂偺怹弫偑擣傔傜傟傑偟偨(恾2B)丅偙偺偙偲偐傜TIARP偼娭愡墛偺梷惂婡峔偵旕忢偵廳梫側摥偒傪偟偰偄傞偙偲偑柧傜偐偲側傝傑偟偨丅 A.
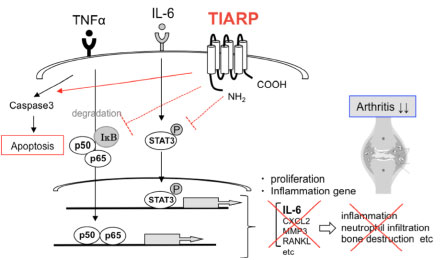
B. 恾2. TIARP寚懝儅僂僗偼CIA昦懺偑憹埆偡傞 亂TIARP偺娭愡墛梷惂婡峔偺夝柧亃 変乆偼TIARP偺娭愡墛惂屼儊僇僯僘儉傪傛傝柧傜偐偵偡傞偨傔偵丄TIARP敪尰嵶朎偱偁傞儅僋儘僼傽乕僕偵拝栚偟偰専摙傪峴偄傑偟偨丅TIARP寚懝M兂偵TNF兛偱巋寖偡傞偲丄IL-6嶻惗検偑TIARP寚懝儅僂僗偱挊柧偵憹壛偟傑偟偨丅傑偨丄偦偺壓棳僔僌僫儖偱偁傞NF-kB偺梷惂場巕偲偟偰偟傜傟傞I內B兛敪尰検偼丄TIARP寚懝儅僋儘僼傽乕僕偱偼挊柧偵I倠B兛偺帩懕偟偰暘夝偝傟偰偄傞偙偲傪柧傜偐偲偟傑偟偨丅偝傜偵TIARP寚懝儅僋儘僼傽乕僕偼丄TNF兛桿摫惈傾億僩乕僔僗嵶朎悢偑尭彮偟丄caspase-3抈敀偺暘夝偼栰惗宆偲斾妑偟偰尭庛偟偰偄傑偟偨丅摨條偵IL-6僔僌僫儖偵梌偊傞塭嬁偵偮偄偰傕専摙偟偨偲偙傠丄TIARP寚懝儅僋儘僼傽乕僕偼IL-6巋寖屻偵STAT3偺儕儞巁壔偑槾恑偟偰偄傞偙偲偑傢偐傝傑偟偨丅偙偺偙偲偐傜丄儅僋儘僼傽乕僕偵偍偄偰TIARP偼傾億僩乕僔僗傪懀恑偟丄NF-kB偍傛傃STAT3僔僌僫儖傪晧偵梷偊傞偙偲偱墛徢傪惂屼偡傞栶妱傪扴偭偰偄傞偙偲偑柧傜偐偲側傝傑偟偨 (恾3)丅
恾3. TIARP偺娭愡墛惂屼儊僇僯僘儉(僔僃乕儅恾)
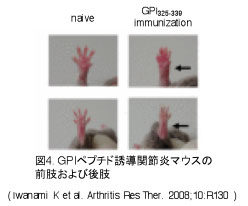
嬃偔傋偒偙偲偵TIARP寚懝儅僂僗偼丄壛楊(12儢寧楊)偵傛傝寣拞IL-6擹搙偺憹壛傪偲傕側偭偰丄娭愡墛傪帺慠敪徢偡傞偙偲偑敾柧偟傑偟偨(恾4)丅偙偺偙偲偼丄TIARP偺寚懝偵傛傝墛徢惈僒僀僩僇僀儞嶻惗偑憹戝偟偨偙偲偵傛傝帺屓柶塽幘姵傪敪徢偡傞偙偲傪帵嵈偡傞傕偺偲峫偊傜傟傑偡丅変乆偺専摙傗丄偙傟傑偱偺抦尒偐傜丄IL-6側偳偺僒僀僩僇僀儞庴梕懱僔僌僫儖堎忢偍傛傃惂屼暘巕偺婡擻堎忢偵傛傝帺屓柶塽幘姵傪敪徢偡傞偙偲傪徹柧偟偨傕偺偱偁傝丄僒僀僩僇僀儞偲帺屓柶塽幘姵敪徢偺婡峔傪峫偊傞忋偱丄旕忢偵廳梫側抦尒偑傕偨傜偝傟傑偟偨丅
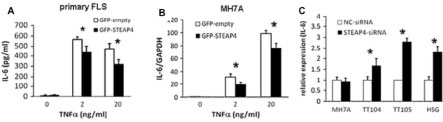
恾4. TIARP寚懝儅僂僗偼娭愡墛傪帺慠敪徢偡傞 尰嵼傢傟傢傟偼丄TIARP偑偳偺傛偆偵偟偰NF-內B傗STAT3側偳偺僔僌僫儖揱払傪惂屼偟偰偄傞偺偐偵偮偄偰専摙傪恑傔偰偄傑偡丅傑偨TIARP婡擻傪槾恑偝傞偨傔偺儕僈儞僪偺摨掕傗峈懱側偳傪嶌惢偡傞偙偲偱丄怴偨側RA帯椕僞乕僎僢僩暘巕偺敪尒傗昦懺惂屼偵岦偗偨怴婯帯椕朄偺奐敪偑恑傓偙偲傪婜懸偟偰偄傑偡丅 亂娭愡儕僂儅僠姵幰偵偍偗傞STEAP4偺婡擻夝愅亃 TIARP偺僸僩ortholog偼丄six-transmembrane epithelial antigen of prostate 4 (STEAP4)偲屇偽傟偰偄傑偡丅STEAP4偼丄慜棫態娻偵崅敪尰偡傞抈敀偲偟偰摨掕偝傟丄崪悜偵師偄偱巕媨傗戀帣娞憻偵敪尰偡傞偙偲偑曬崘偝傟偰偄傑偡丅変乆偺専摙偵傛傝STEAP4偼RA姵幰PBMC偍傛傃妸枌丄摿偵CD68梲惈妸枌嵶朎偵嫮敪尰偡傞偙偲傪尒偄偩偟傑偟偨乮恾5乯丅
恾5. STEAP4偼CD68梲惈RA姵幰妸枌嵶朎偵敪尰偡傞 妸枌嵶朎偵偍偗傞STEAP4偺婡擻傪柧傜偐偲偡傞偨傔偵丄RA姵幰桼棃妸枌嵶朎傊偺STEAP4堚揱巕摫擖偍傛傃僲僢僋僟僂儞傪峴偄傑偟偨丅STEAP4夁忚敪尰妸枌嵶朎偼丄僐儞僩儘乕儖偲斾妑偟偰嵶朎憹怋偑梷惂偝傟丄傾億僩乕僔僗嵶朎偺桳堄側憹壛傪擣傔傑偟偨丅偝傜偵丄TNF兛巋寖埶懚揑側IL-6偍傛傃IL-8嶻惗傪梷惂偡傞偙偲偑柧傜偐偲側傝傑偟偨丅
恾6. STEAP4偼丄IL-6嶻惗傪梷惂偡傞 偙偺暘巕偺僸僩偵偍偗傞堄媊偵偮偄偰偼枹偩晄柧側揰傕悢懡偔丄RA姵幰妸枌嵶朎偺傎偐寬忢恖枛徑寣偺岲拞媴側傜傃偵扨媴偵傕敪尰偑擣傔傜傟傞偙偲偐傜丄奺乆偺嵶朎偵偍偗傞STEAP4暘巕偺婡擻夝愅側偳傪拞怱偵丄尰嵼専摙傪恑傔偰偄傑偡丅
|
|