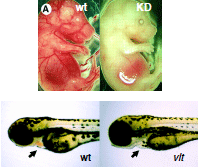
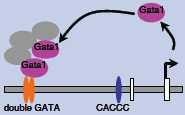
|
【参考文献】
1. Kobayashi and Yamamoto (2005) Molecular mechanisms activating the Nrf2-Keap1 pathway of antioxidant gene regulation. Antioxid. Redox Signal., 7: 385-394.
2. Suzuki et al. (2005) Pi-class glutathione S-transferase genes are regulated by Nrf2 through an evolutionarily conserved regulatory element in zebrafish. Biochem. J., 388: 65-73.
3. Takagi et al. (2004) MafT, a new member of the small Maf protein family in zebrafish. Biochem. Biophys. Res. Commun., 320: 62-69.
4. Kobayashi et al. (2002) Identification of the interactive interface and phylogenic conservation of the Nrf2-Keap1 system. Genes Cells, 7: 807-820.
5. 小林麻己人(2002)親電子性物質センサー分子同定を目指したゼブラフィッシュの活用 実験医学 20: 1656-1661.
|