|
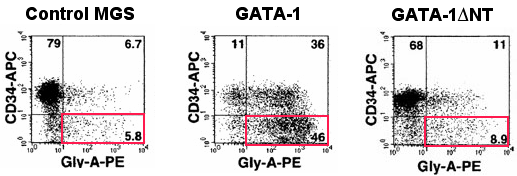
傑偨丄変乆偼Complementation Rescue朄傪梡偄偰丄GATA-1傪捠忢偺5亾掱搙偟偐敪尰偣偢丄GATA-1儮NT傪僩儔儞僗僕乕儞乮TG乯偲偟偰帩偮丄GATA-1.05:: GATA-1儮NT TG儅僂僗傪嶌惢偟丄GATA-1儮NT偑寣媴暘壔偵傕偨傜偡婡擻傪屄懱儗儀儖偱専徹偟偨丅GATA-1儮NT掅敪尰儔僀儞偱偼愒寣媴暘壔傪桿摫偡傞偙偲偼偱偒側偄偑丄崅敪尰儔僀儞偱偼愒寣媴暘壔傪桿摫偡傞偙偲偑偱偒偨丅偙傟傜偺偙偲偐傜丄GATA-1偺N枛抂偑寣媴暘壔偵懳偟偰廳梫側婡擻傪帩偭偰偄傞偙偲偑傢偐傝丄尰嵼丄偦偺暘巕儊僇僯僘儉傪夝愅偟偰偄傞偲偙傠偱偁傞丅
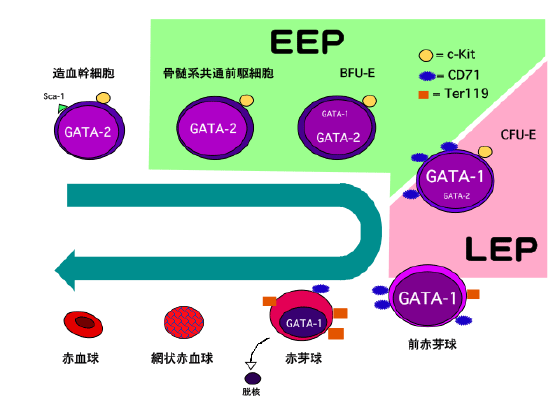
僟僂儞徢偱側偤AMKL偑崅棪偵敪徢偡傞偺偐丄偦偺儊僇僯僘儉偼傑偩姰慡偵夝柧偝傟偨傢偗偱偼側偄丅GATA-1偺N枛抂偑寚幐偟偰偄傞偙偲丄僟僂儞徢偱偼俀侾斣愼怓懱偑俁杮偁傞偙偲偑娭梌偟偰偄傞偺偱偼側偄偐偲悇掕偝傟傞丅偦偙偱丄21斣愼怓懱忋偵偁傝丄寣媴偵娭楢偑偁傞揮幨場巕RUNX1傗Bach1偑岓曗偲側傝丄尋媶偑恑傔傜傟偰偄傞丅
丂変乆偼丄GATA-1 N枛抂偺寣媴暘壔丒憹怋偵偍偗傞婡擻傪夝偒柧偐偡傋偔丄暘巕惗暔妛揑庤朄偲儅僂僗敪惗岺妛揑庤朄傪梡偄偰丄帋尡娗儗儀儖偲屄懱儗儀儖偺椉柺偐傜傾僾儘乕僠偟偰偄傞偲偙傠偱偁傞丅
|