|
岩坪 威、高杉展正、富田泰輔(東京大学・大学院薬学系研究科・臨床薬学教室)
常染色体優性遺伝を示す早発型家族性アルツハイマー病(familial
AD;FAD)の主要な病因遺伝子として1995年に同定されたプレセニリン(presenilin;PS)の変異が、凝集性が高く早期からAD脳に蓄積するAβ421)の産生を上昇させることが96-97年にかけて明らかになり2)、PSのFAD効果はAβのC末端形成−γ切断−に関連することがコンセンサスとなった。次に問題になったのはPSとγセクレターゼの関係である。
γセクレターゼはAPP, Notchなどの一回膜貫通型蛋白の膜貫通領域内での切断(intramembrane proteolysis)を担うユニークなプロテアーゼである。PS1ノックアウトマウス由来の細胞においてγセクレターゼ活性が高度に低下したことから、PSはγ活性に不可欠の分子であることがわかった3)。さらにPSの膜貫通領域内の2個のアスパラギン酸
(PS1ではAsp257とAsp385)に変異を導入するとγセクレターゼ活性が消失すること4)、アスパルチルプロテアーゼの活性中心に結合する「遷移状態模倣型阻害剤」がγセクレターゼを抑制するとともに断片型のPSに結合することが示され5)、PSは非定型的な膜貫通型プロテアーゼであるとの見解がにわかに有力になった。さらにPS単独ではγセクレターゼ活性は生じないが、断片型PSと他の蛋白性構成因子(co-factors)が250~1000
kDa以上の高分子量の複合体を形成して、γセクレターゼ活性を発揮することが明らかになった6)。これらの知見から、現在γセクレターゼは膜結合型の巨大なアスパラギン酸性プロテアーゼ複合体であり(あたかもプロテアソームを想起させる点で、membrasome,
secretosomeなどの造語も生まれた)、PSはその活性中心を形成する触媒サブユニットと捉えられるようになった。このような経緯から、PS以外のγセクレターゼ複合体の構成タンパク質(cofactor)の同定がγセクレターゼ研究の焦点となった。
γセクレターゼの必須のパートナー蛋白として最初に同定されたのはニカストリン (nicastrin; NCT)である7)。西村正樹とYuらはジギトニンで可溶化した膜画分中のPS複合体を抗PS抗体を用いてイムノアフィニティ精製し、I型の1回膜貫通蛋白NCTを同定した。ヒトNCTは約700アミノ酸からなる糖蛋白であり、機能型γセクレターゼ複合体に含まれるNCTは複合型N結合型糖鎖付加を受けている8)(しかし糖付加は活性に必要ではないらしい)。Yuらは線虫のNCTホモログaph-2遺伝子をRNAi法でノックダウンすると、線虫notchホモログの一つであるglp-1欠損変異に類似した咽頭前部の低形成(anterior pharynx hypoplasia)を生じることを示し、NCTがPSと同様にnotch活性化に必須であることを示した。またStruhl9),
Fortini 10)らはショウジョウバエNCTを欠損させると、notchシグナルの障害により胚致死となるが、この表現型はγセクレターゼによるnotch切断産物であるNICDによりrescueされること、NCT変異個体やNCTをRNAiでノックダウンしたショウジョウバエ細胞ではNICDが産生されず、機能型のPS断片も消失することを示し、NCTがγセクレターゼ複合体の形成とその機能に必須の因子であることを実証した。しかしNCTの過剰発現のみでは機能型のPS複合体が増量しないことから10)、γセクレターゼの量や活性を決定するcofactorが他にも存在するものと考えられた。
2002年1月、米国のGoutteらは、機能欠損により線虫NCT (aph-2)11)の変異に類似した咽頭部発生異常を示す遺伝子aph-1を同定した12)。Aph-1は二百数十アミノ酸からなる7回膜貫通型の膜蛋白をコードしており、表現型から考えるとγセクレターゼ複合体の新しい構成因子である可能性が高いと考えられた。こうして2002年の初頭からγセクレターゼ研究者の間では、密かに「第3ラウンド」が始まっていた。
2002年6月26日のこと、筆者の研究室でPS複合体の研究に取り組んでいた高杉らは、インターネットでaph-1について情報検索すると、pen-2という見慣れぬ遺伝子の記載があるのに気づいた。南サンフランシスコのベンチャー企業Exelixis社が、4月の米国ショウジョウバエ学会で、新たなγセクレターゼcofactor候補分子としてpen-2を発表した記録がアップロードされていたのだった。この時点でデータベースを検索すると、ショウジョウバエやヒトのpen-2の遺伝子情報も入手することができた。ショウジョウバエS2細胞においてAPH-1をRNAi
ノックダウンするとγセクレターゼ活性が消失するというデータをすでに得ていた高杉らは、PEN-2もγセクレターゼの必須因子に違いないと確信し、ただちにRNAiや過剰発現実験に着手した。翌週末の7月5日、APH-1,
PEN-2に関する論文は今週も出なかったと胸をなで下ろそうとした矢先、Exelixis社の膨大な研究成果がDevelopmental Cell誌に公表された13)。Dan
Curtisの率いる研究チームは線虫のnotch表現型の増強、抑制を指標とする系統的な遺伝子スクリーニングを行い、PSによるγセクレターゼ機能を活性化する遺伝子
(presenilin enhancer)としてpen-1, pen-2を同定した。Pen-1はaph-1に一致したが、pen-2は新規の2回膜貫通蛋白をコードしていた。Curtisらも、ショウジョウバエS2細胞を用いて内因性のaph-1,
pen-2をRNAiでノックダウンすると、断片型ショウジョウバエPS (Psn)が消失するとともに、共発現したヒトAPP C末端断片からのAβの産生が抑制されるという結果を記載していた。これは高杉のAPH-1に関するデータとあまりにも一致しており、我々は狼狽した。この時点でAPH-1,
PEN-2がともにγセクレターゼ複合体の形成、機能に必須な新たなPS cofactorであることが明らかになったわけであるが、NCT, APH-1, PEN-2が個別に果たす役割はまだ不明であった。
ちなみにアルツハイマー研究者の中でcofactorレースに最も早く乗り出していたのは、ボストンのSelkoeらであった。同グループのMike Wolfeは、γセクレターゼ阻害剤をショウジョウバエ個体に適用するとnotch表現型が生じるという結果を4月のショウジョウバエ学会で自ら発表した際、Curtisらの発表を目の当たりにした。そこでSelkoeはただちにCurtisをセミナーに招き、APH-1,
PEN-2の研究に着手したとのことである。
富田、高杉らは、PSが昆虫や無脊椎動物に至るまで高度に保存された分子であることに注目し、RNAi法によるノックダウンが最初に可能となっていたショウジョウバエS2細胞を利用してγセクレターゼ複合体を解析しようと着想した(2000年の盛夏、当時阪大岡野研の助教授であった三浦正幸博士のもとにS2細胞を頂きに行ったころ、まだ哺乳類細胞でsmall
interfering RNAは利用可能になっていなかった)。高杉らは、まずショウジョウバエAPH-1 (dAPH-1)とNCT (dNCT)をS2細胞に恒常発現し、ショウジョウバエPS
(Psn)代謝の変化を調べた。するとdNCTを発現しただけではPsnの代謝に変化は生じないが、dAPH-1を発現した場合、断片型Psnの量は変わらないのに対し、全長型Psnが増量することに気づいた。dAPH-1とdNCTを共発現すると、全長型Psnはさらに増量した。この全長型Psnは断片型と同様250
kDa以上の高分子量複合体を形成しており、高度に安定化されていたが、この両者の共発現のみではγセクレターゼ活性は増大しなかった。次にショウジョウバエPEN-2
(dPEN-2)をRNAiでノックダウンすると、断片型Psnが消失すると同時に、dNCTやdAPH-1を抑制した場合と異なり、全長型Psnが蓄積してくるのに気づいた。前述のCurtisらの論文では、同じくS2細胞を用いたdPEN-2のRNAiノックダウンによって、Psnは断片型、全長型ともに高度に減少したデータが記載されている。しかしCurtisらは6回のRNAi実験のうち1回のみではあるが、全長Psnの蓄積を観察していたとのことである(私信)。この全長型Psnは高分子量複合体を形成し、高度に安定化されていた点で、dAPH-1,
dNCT共発現の場合ときわめて似た性質を示した。そしてdAPH-1, dNCTに加えてdPEN-2の3者を共発現することにより、断片型Psnの産生と、γセクレターゼ活性の増大が確認された。これらの結果から、我々は次のように結論した(図1)。(1)APH-1はγセクレターゼ複合体の主要な安定化因子として作用する。(2)PEN-2はγセクレターゼ複合体活性化の最終段階を担い、安定化したPSにγセクレターゼ活性を与えるとともに、PSの断片化を促す。(3)PS,
NCT, APH-1, PEN-2は、γセクレターゼ複合体の基本骨格を構成する主要な構成因子である14)。
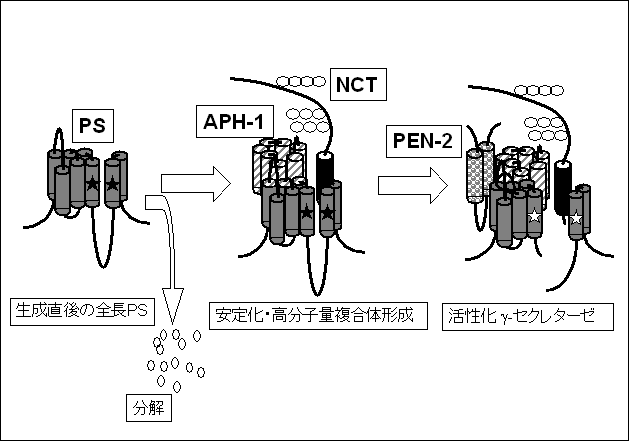 |
図1 γセクレターゼ複合体の形成過程に関する仮説。
生成されたばかりのPSの大部分は速やかに分解され、低分子量にとどまる(左)。PSの一部はAPH-1およびNCTと結合し、高度に安定化された高分子量の複合体を形成するが、この時点ではPSはプロセシングを受けていない全長蛋白の状態で存在し、γセクレターゼ活性もない(★で非活性型のAsp残基を表示;中)。引き続いてPEN-2が結合することによりγセクレターゼの活性化が起こり(☆で活性型のAsp残基を表示)、PSのプロセシングも生じる(右)。
|
しかし未解決の問題がまだ多く残されている。我々はPSに加えて3種類のcofactorを共発現することによりγセクレターゼ複合体の形成と活性化が生じることを確認したが、断片型PSの増加量に見合うだけのγ活性の増大が見られない傾向がある。γセクレターゼの活性化には、単純に4者が共発現するのみでは不十分であり、他に活性化のメカニズムを想定すべきなのかもしれない。またこの4者の分子量を単純に合計しただけでは200
kDaに満たず、第5の構成因子が存在する可能性も十分に残される。複合体中にこれらの因子がどのような量比で含まれているのかも重要な問題である。さらにPEN-2がいかなる分子機構によって複合体を活性化するのか、PSの膜貫通部位に存在する2個のアスパラギン酸残基が、どのような機序で膜内に埋まった基質の一定位置に加水分解--γ切断--を及ぼしうるのか、そしてPSの1アミノ酸置換がどうやってAβ42ペプチドの産生を招くのか、など課題は尽きない。Haassらはごく最近、内因性にPSを持たない酵母細胞にPSを含む4者を共発現すると、γセクレターゼ活性が生じることを報告した。しかしγセクレターゼとPS複合体の同一性を厳密に証明するためには、個別に調製されたリコンビナント蛋白を混和して、γセクレターゼ活性を試験管内で再現するという形での完全なin
vitro再構成が求められよう。また活性型のγセクレターゼ複合体を結晶化し、立体構造解析を行うことも将来的な重要課題であろう。
家族性ADの病因遺伝子として同定されたPSを出発点として、γセクレターゼ複合体による「膜内蛋白質分解機構」の研究は、Notchなどの発生・分化に関連した重要因子の研究と合流し、この数年間に魅力的な研究テーマとなった。また、Notch切断抑制などの副作用の懸念は残されるにしても、γセクレターゼ阻害剤は依然としてADの根本治療薬の有力な候補の一つである。今後の展開が楽しみである。
文献
1.Iwatsubo T, Odaka A, Suzuki N, Mizusawa H, Nukina
N, Ihara Y. Visualization of Aβ42(43) and Aβ40 in senile plaques with
end-specific Aβ monoclonals: evidence that an initially deposited species
is Aβ42(43). Neuron 13: 45-53, 1994
2.Tomita T, Maruyama K, Saido TC, Kume H, Shinozaki
K, Tokuhiro S, Capell A, Walter J, Grunberg J, Haass C at el. The presenilin
2 mutation (N141I) linked to familial Alzheimer disease (Volga German
families) increases the secretion of amyloid β protein ending at the
42nd (or 43rd) residue. Proc Natl Acad Sci USA 94:2025-2030, 1997
3.De Strooper, B., Saftig P., Craessaerts, K., Vanderstichele,
H., Guhde, G., Annaert, W., Von Figura, K. and Van Leuven, F. Deficiency
of presenilin-1 inhibits the normal cleavage of amyloid precursor protein.
Nature 391: 387-390, 1998
4.Wolfe MS, Xia W, Ostaszewski BL, Diehl TS, Kimberly
WT, Selkoe DJ. Two transmembrane aspartates in presenilin-1 required
for presenilin endoproteolysis and γ-secretase activity. Nature 398:
513-517, 1999
5.Li YM, Xu M, Lai MT, Huang Q, Castro JL, DiMuzio-Mower
J, Harrison T, Lellis C, Nadin A, Neduvelil JG et al. Photoactivated
γ-secretase inhibitors directed to the active site covalently label
presenilin 1. Nature 405: 689-694, 2000
6.Tomita T, Watabiki T, Takikawa R, Morohashi Y, Takasugi
N, Kopan R, De Strooper B, Iwatsubo T. The first proline of PALP motif
at the C terminus of presenilins is obligatory for stabilization, complex
formation and γ-secretase activities of presenilins. J Biol Chem 276:33273-33281,
2001
7.Yu G, Nishimura M, Arawaka S, Levitan D, Zhang L,
Tandon A, Song YQ, Rogaeva E, Chen F, Kawarai T et al. Nicastrin modulates
presenilin-mediated notch/glp-1 signal transduction and βAPP processing.
Nature 407:48-54, 2000
8.Tomita T, Katayama R, Takikawa R, Iwatsubo T. Complex N-glycosylated
form of nicastrin is stabilized and selectively bound to presenilin
fragments. FEBS lett 520:117-121, 2002
9.Chung HM, Struhl G. Nicastrin is required for presenilin-mediated
transmembrane cleavage in Drosophila. Nat Cell Biol 3:1129-1132,
2001
10.Hu, Y., Ye, Y. Fortini, M.E. Nicastrin is
required for g-secretase cleavage of the Drosophila Notch receptor.
Dev Cell 2: 69-78, 2002
11.Goutte C, Hepler W, Mickey KM, et al. aph-2 encodes
a novel extracellular protein required for GLP-1-mediated signaling.
Development 127:2481-2492, 2000
12.Goutte C, Tsunozaki M, Hale VA, Price JR. APH-1
is a multipass membrane protein essential for the Notch signaling pathway
in Caenorhabditis elegans embryos. Proc Natl Acad Sci USA. 99: 775-779,
2002
13.Francis R, McGrath G, Zhang J, Ruddy DA, Sym M,
Apfeld J, Nicoll M, Maxwell M, Hai B, Ellis MC et al. aph-1 and pen-2
are required for Notch pathway signaling, γ-secretase cleavage of βAPP,
and presenilin protein accumulation. Dev Cell 3:85-97, 2002
14.Takasugi N, Tomita T, Hayashi I, Tsuruoka M, Niimura
M, Takahashi Y, Thinakaran G,Iwatsubo T. The roles of presenilin cofactors
in the γ-secretase complex. Nature 422:438-441, 2003
15.Edbauer D, Winkler E, Regula JT, Pesold B, Steiner
H, Haass C. Reconstitution of γ-secretase activity. Nat Cell Biol 5:486-488,
2003
|
