西村隆史、貝淵弘三 (名古屋大学・大学院医学系研究科・細胞情報薬理学講座)
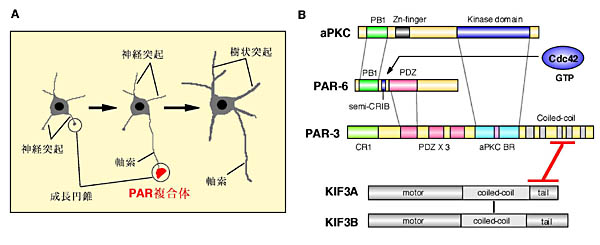
神経細胞は、軸索と樹状突起という形態的にも機能的にも異なる突起を持っている(図1A)。樹状突起は他の神経細胞から情報を受け取り、軸索はその情報を他の細胞に伝達する。神経細胞には樹状突起から軸索へという方向性を持ったシグナルの流れが存在している。しかし、神経細胞が極性を獲得、維持していくメカニズムには不明な点が多く残されている1-3)。我々は、神経細胞の極性形成の分子メカニズムを解明するために、PAR-3蛋白質に着目した。
PAR-3はPAR-6およびatypical PKC(aPKC)と共に、様々な生物・細胞種において細胞極性形成に重要であると考えられている分子である4-6)。PAR-3は3つのPDZ
ドメインおよびaPKCと結合する領域をもっており、PAR-3、PAR-6およびaPKCはお互いに直接結合する(図1B)。また、PAR-6は極性形成に関与する低分子量GTP結合蛋白質RhoファミリーのCdc42と結合し、PAR-3、PAR-6、aPKCおよびCdc42の四者が協調して極性形成に関わるものと考えられている。例えば、線虫においてPar-3は線虫の受精卵の中で非対称に分布し、Par-6とPKC-3(aPKC)と結合して受精卵の非対称分裂に重要な役割を果たしている。また、ショウジョウバエにおいてPAR-3のホモログであるBazookaの機能を阻害すると、胚上皮のadherens
junctionの形成不全や、胚上皮や神経芽細胞の紡錘体の方向が異常になることから非対称分裂に重要な役割を果たすことが知られている。一方、哺乳類上皮細胞においてPAR-3はaPKCと共にtight
junctionに局在し、管腔側と基底膜側の極性形成に重要であることが知られている。最近、培養海馬神経細胞の極性形成においても、PAR複合体が関与することが報告された7)。PAR-3及びPAR-6は軸索先端の成長円錐に濃縮した局在を示す。PAR-3、PAR-6の過剰発現およびaPKCの阻害剤を用いた実験より、PAR複合体の軸索への局在およびaPKCの活性化が軸索決定に必要であることが示された。しかし、細胞極性形成におけるPAR複合体の機能は、その特徴的な局在に依存しているものの、神経細胞を含めPAR複合体の局在化機構は不明であった。
|
図1.神経細胞の極性形成機構とPAR複合体
A)海馬より取り出してきた神経細胞を培養すると、4〜5本の短い神経突起が細胞体から伸長する。培養1〜1.5日たつと、神経突起のうちの1本が急速に伸長し、形態上の極性(軸索)が観察されるようになる。PAR複合体は軸索先端の成長円錐に濃縮した局在を示す。B)PAR複合体(PAR-3,
PAR-6, aPKC)、低分子量GTP結合蛋白質Cdc42、およびKIF3モーターの結合様式。 |
哺乳類のPAR-3には分子量が100kDa、150kDa、180kDaの3つのスプライシングバリアントが存在しており、組織特異的な発現パターンが知られている。脳には180kDaのPAR-3が多く発現している。そこで我々は、180kDa特異的領域を含むフラグメントを用いてyeast
two hybrid法によるPAR-3結合蛋白質の探索を行ったところ、KIF3Aを同定した8)。KIF3Aは、KIF3BまたはKIF3Cとヘテロ二量体を形成して、微小管のプラス端に向かって動く微小管依存性モータータンパク質である。また、KIF3モーターは、神経組織に高レベルに発現しており、軸索輸送に関与することが知られている。In
vitro の結合実験より、PAR-3はKIF3AのC末端領域に直接結合することが確認された(図1B)。またPAR-3はKIF3Bには結合しなかった。PAR-3のKIF3A結合領域は、PAR-3のC末端側180kDa特異的領域に同定された。よって、PAR-3はKIF3Aに特異的に直接結合することが明らかとなった。また、ラット胎児脳およびCOS7培養細胞を用いた免疫沈降実験より、PAR-3-aPKC複合体がKIF3モーターと生理的に結合していることが確認された。
次に、初代培養海馬神経細胞において、PAR-3およびKIF3の局在を検討したところ、軸索先端部の成長円錐において共局在が観察された。また軸索のシャフトにおいても共局在が確認された。更に、aPKCも軸索先端部の成長円錐に濃縮した局在を示すことが明らかとなった。よって、PAR-3-aPKC複合体はKIF3モーターによって軸索の先端まで運ばれている可能性が考えられた。この可能性を検討するために、PAR-3およびKIF3の様々なドミナントネガティブ変異体を作製した。まず、PAR-3とKIF3Aの結合領域断片に着目した。PAR-3とKIF3Aの結合領域断片を過剰発現させると、PAR-3とKIF3Aの結合が阻害される可能性が考えられる。実際にin
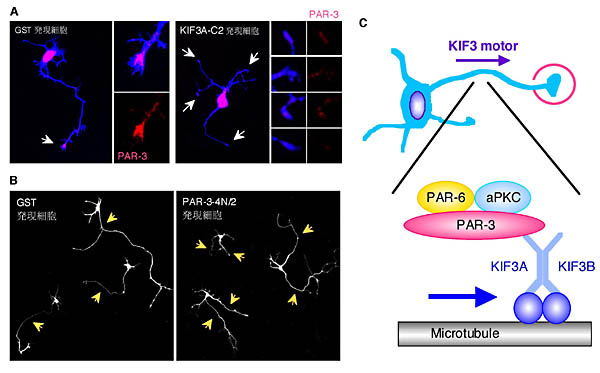
vitro において、PAR-3および KIF3Aの結合領域断片は、量依存的にPAR-3とKIF3Aの結合を阻害した。これら断片を神経細胞に過剰発現させたところ、PAR-3およびaPKCの神経突起先端部への局在を阻害した(図2A)。さらに、KIF3AまたはKIF3Bのモーター部分を欠損した変異体を用いて実験を行った。KIF3モーターはヘテロ2量体を形成して動くため、3Aまたは3Bのモーター部の欠損変異体はKIF3モーター活性を阻害すると考えられる。これら変異体は、同様にPAR-3およびaPKCの神経突起先端部への局在を阻害した。以上の結果より、PAR-3-aPKCはKIF3モーターによって神経突起先端部まで運ばれていることが示唆された。
|
図2.神経細胞におけるPAR複合体の局在化と極性形成
A)GST(コントロール)およびKIF3A-C2(KIF3AのPAR-3結合領域フラグメント)発現細胞における内在性PAR-3の局在。神経突起先端部(矢印)の拡大図をそれぞれ横に示す。B)GST(コントロール)およびPAR-3-4N/2(PAR-3のKIF3A結合領域フラグメント)発現細胞の形態。コントロール細胞では1本の長い軸索が形成されるのに対し、PAR-3-4N/2発現細胞では1本の長い軸索が形成されず、同様の長さの神経突起が形成された。C)神経細胞におけるPAR複合体局在化機構のモデル。(文献8より改変)
|
PAR-3およびKIF3のドミナントネガティブ変異体により、PAR-3またはaPKCの局在異常を起こしている細胞の多くは、軸索形成の異常も観察された(図2B)。海馬より取り出してきた神経細胞を培養すると、4〜5本の短い神経突起が細胞体から伸長する(ステージ2)1-3)。この段階の神経突起は、形態的にはお互いにほぼ等価であると考えられている。培養1〜1.5日たつと、神経突起のうちの1本が急速に伸長し、形態上の極性が観察されるようになる(ステージ3)。この急速に伸長した神経突起は軸索としての性質を有するようになる。PAR-3およびKIF3のドミナントネガティブ変異体を発現している細胞は、1本の長い軸索が形成されず、複数の短い軸索様突起を形成していた。以上の結果から、PAR-3-aPKCの複合体がKIF3モーターによって神経突起先端部に運ばれ、局在化することが神経細胞の極性形成(軸索形成)に重要であることが示唆される(図2C)。
このようにPAR複合体は神経突起先端部に局在化することで、神経突起の伸長および軸索形成を制御していると考えられる。また、PI3キナーゼ活性もPAR-3の神経突起先端への局在に関与している7)。PI3キナーゼとRhoファミリータンパク質の局所的な活性化は細胞極性形成のシグナルの上流に位置すると考えられている。実際に、神経細胞において神経突起の伸長に先立ち、神経突起のうちの1本でPI3キナーゼの産物であるPI(3,4,5)P3が濃縮する9)。よって、細胞外基質からPI3キナーゼ、さらにCdc42のシグナル伝達系がPAR複合体の上流シグナルとして軸索形成に関与していると考えられる7),10)。また、PAR複合体の下流シグナルとして、我々はPAR-3がRac特異的活性化因子であるSTEF/Tiam1と結合し、Racの活性化に関与することを見出している(西村ら、投稿中)。今後は、PAR-3の上流および下流シグナルを明らかにしていくことで神経細胞の極性形成の分子メカニズムが解明されていくのではないかと考えている。
謝辞
本研究は、横浜市立大学医学部の大野茂男教授との共同研究として行ったものです。記して深謝いたします。
文献
1) Craig, A. M. & Banker, G. : Neuronal polarity. Annu. Rev. Neurosci.
17 : 267-310, 1994
2) Fukata, Y. et al. : Axon specification in hippocampal neurons. Neurosci.
Res. 43 : 305-315, 2002
3) 西村隆史ら : 神経細胞の極性形成機構, 実験医学 21 : 2324-2329, 2003
4) Macara, I.G. : Parsing the polarity code. Nature Rev. Mol. Cell Biol.
5 : 220-231, 2004
5) Ohno, S. : Intercellular junctions and cellular polarity: the PAR-aPKC
complex, a conserved core cassette playing fundamental roles in cell
polarity. Curr. Opin. Cell Biol. 13 : 641-648, 2001
6) Jan, Y. N. & Jan, L. Y. : Asymmetric cell division in the Drosophila
nervous system. Nature Rev. Neurosci. 2 : 772-779, 2001
7) Shi, S. H. et al. : Hippocampal neuronal polarity specified by spatially
localized mPar3/mPar6 and PI 3-kinase activity. Cell 112 : 63-75, 2003
8) Nishimura, T. et al. : Role of the PAR-3-KIF3 complex in the establishment
of neuronal polarity. Nature Cell Biol. 6 : 328-334, 2004
9) Menager, C. et al. : PIP3 is involved in neuronal polarization and
axon formation. J. Neurochem. 89 : 109-118, 2004
10) Arimura, N. et al. : Role of CRMP-2 in neuronal polarity. J. Neurobiol.
58 : 34-47, 2004
|