松崎 政紀 (自然科学研究機構・生理学研究所)
1.研究の背景
脳の学習・記憶の素過程はシナプスの伝達効率の変化(シナプス可塑性)にあると考えられている。特に、シナプス前線維とシナプス後細胞が同時に頻回発火した時にシナプス伝達効率が増加する長期増強(LTP)の現象は、記憶形成に関与するとされる。最近の研究結果から、シナプス前終末から放出されたグルタミン酸を受容するシナプス後部AMPA受容体の発現量が変化(LTP時には増加)することが、シナプス可塑性の要因として最重要視されている。だが少数の代謝回転が早い分子だけで、長期にわたる記憶を保持できるとは想像しにくい。もしシナプス構造自身がシナプス可塑性時に変化して固定すれば、構造はそこに存在するすべての分子に影響を与えるため、シナプス伝達効率に関与する分子群の構成を安定して維持できると推測される1)。
AMPA受容体が発現しているシナプスの後部は、興奮性神経細胞においては主にスパイン(spine、棘突起)と呼ばれる構造体に作られる。スパインは、1-2
個/μmの密な間隔で樹状突起から突き出しており、ひとつの細胞あたり数千から数万個存在する。スパインは頭部(体積0.004-0.56 μm3)と、頭部と樹状突起の幹部をつなぐネック(直径0.04-0.5
μm、長さ0.08-2 μm)から成る2)。スパインは1
μm3以下の微細な構造ではあるが、頭部体積でいえば、最大のものと最小のもので100倍以上の差がある程、その形には多様性がある。この形態の多様性がシナプス可塑性を反映しているかどうかについては数多く議論されてきた3)。しかし例えば電子顕微鏡による観察では、多数のスパインの中からLTPが起きたスパインを完全に同定することができず、一致した結論を導くことができていない。
1990年代になって2光子励起顕微鏡が登場すると、脳スライス標本中のスパイン形態、スパイン内カルシウム動態を生きたままイメージングすることが可能となった。その結果、スパインのネックはカルシウムイオン等の代謝因子をスパイン内に遮蔽する効果を持つことがわかり、スパインが個別に機能している可能性が示唆された。それならば、記憶は個々のスパインを最小単位として蓄えられているのだろうか。しかしながらいまだに、同定された1個のスパインでLTPを誘発した実験は報告されていない。狙ったスパインに入力するシナプス前線維1本だけをテタヌス刺激し、同時にスパイン形態と機能変化(AMPA受容体反応の変化)を観察することが極めて困難だからである。我々は本研究において、2光子励起顕微鏡という技術を刺激方法に応用することによって、単一スパインで、スパイン形態の変化とAMPA受容体反応の変化を同時に捉えることに成功したので、ここに報告する。
2.研究の概要
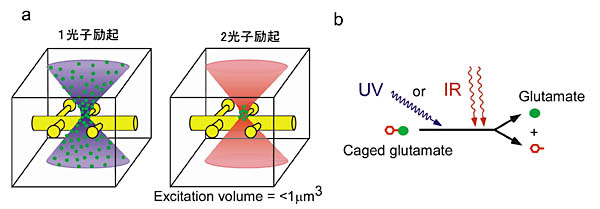
近赤外超短光パルスレーザーをレンズを用いて集光させると、焦点領域での光密度が顕著に高くなり、可視光付近に吸収帯を持つ分子に対し、2つの光子をほぼ同時に吸収させ、励起させることができる(図1a)。光子を吸収する分子が蛍光分子ならば、焦点領域のみから光が放射されるので、断層効果をもった蛍光画像を得ることができる。これが2光子励起蛍光イメージングである。そこで我々は、この方法をケイジドグルタミン酸に適用することを試みてきた。水溶液中に溶解したケイジドグルタミン酸は、レーザー焦点領域でのみ2光子を吸収して、分子内解離反応を起こし、グルタミン酸を放出する(図1b)。焦点領域は1
μm3以下なので、微細な構造であるスパインに対し、単一スパインレベルで、グルタミン酸を投与することができるという原理である。光で刺激するので、スライス標本のような組織内に深く埋まっているスパインにも適用可能である。さらに重要な利点は、シナプス小胞の放出確率や小胞内グルタミン酸量のゆらぎ等の影響を受けることなしに、スパイン表面で機能しているAMPA受容体、NMDA受容体の反応を誘発できることにある。
|
図1
(a) 1光子励起では光が通るすべての領域で分子が励起されるが、2光子励起では焦点領域でのみ分子が励起される。(b) ケイジドグルタミン酸は紫外光子1個を吸収するとグルタミン酸を放出する。このエネルギーに相当する近赤外光子を2個ほぼ同時に吸収した場合にも、グルタミン酸を放出できる。 |
まず本研究の前段階として、我々は単一スパイン形態とAMPA受容体発現の関係性を調べた4)。海馬スライスのCA1錐体細胞にホールセルパッチクランプを行い、樹状突起を含むある小領域すべてにおいて、MNI-glutamateという新しいケイジドグルタミン酸を2光子励起法によって活性化し、そのときのAMPA受容体反応を記録した。その結果、2光子励起法によるケイジドグルタミン酸の活性化という方法論によって、単一スパインのAMPA受容体反応が実際に得られることがわかった。さらに、スパイン頭部体積とAMPA受容体の機能的発現は、強く正に相関しており、大きなスパインほど多くのAMPA受容体が機能していることが判明した。
それではLTP時には、スパイン形態はどうなっているだろうか。上記の結果からは、AMPA受容体発現が増えれば、いずれは頭部も大きくなると予想される。我々の方法を用いれば、同定された単一スパインに対してLTPを誘発できるかもしれない。しかしそもそも、シナプス前終末の活動なしに、グルタミン酸を投与しただけで、LTPは起こるのだろうか。
これらの問題に取り組むためには、スパイン形態を長時間観察しながら、2光子励起法によって刺激をする必要がある。そこで、我々は2光子励起顕微鏡をさらに改良して、ひとつの顕微鏡に2つのレーザーを独立した走査ユニットを通して導入した。この顕微鏡では、720
nmの波長のレーザーを用いて、標本中のある一点を照射してケイジドグルタミン酸を活性化し、同時に、もうひとつの910 nmのレーザーによって蛍光観察をすることができる。スパイン形態は、神経細胞に発現させたGFPによって観察し、なるべく生体内に近い状態で調べるため、海馬スライス培養標本を用いた。GFP遺伝子は遺伝子銃を用いて細胞に導入した。GFPを発現したCA1錐体細胞の樹状突起スパインを任意にひとつ選択し、2光子励起法によって刺激した。細胞外からNMDA受容体を阻害するマグネシウムを除き、テトロドトキシンを入れた状態で、1
Hz 60秒の刺激を行った。一回の刺激で放出されるグルタミン酸量は、パッチクランプしている状態で調べると、mEPSCとほぼ同じ大きさの電流反応(<25
pA)を誘発することから、一個のシナプス小胞内のグルタミン酸量におおよそ相当する。
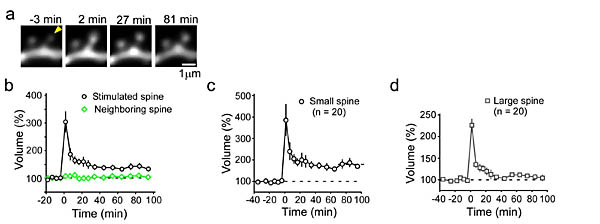
そうすると、刺激されたスパインに選択的にその頭部体積が、照射10秒以内に増加し始め、5分以内に最大値300%に達した(図2)。30%程度のスパインは、20分後には150%増大まで戻った後、その大きさが100分以上維持された。この時間経過は、電気刺激で観察される長期増強の形成パターンとほぼ一致する。一過性の体積増大は、小さなスパイン(0.1
μm3未満)では100%観察され、0.1 μm3より大きくなると90%で観察された。しかし、持続性の体積増大は小さいスパインでは55%で観察されたが、大きいスパインでは5%にしか見られなくなった。100
Hz 1秒のシナプス前線維に対する電気刺激では、小型スパインのみに持続的な体積増大が生じた。この形態可塑性は、NMDA受容体阻害剤、アクチン重合阻害剤、カルモジュリン阻害剤によってほぼ完全に阻害された。これらはいずれも、LTPを抑制することが知られている。一方で、CaMキナーゼIIの阻害剤は一過的な増大を阻害しなかったが、体積増大維持を阻害した。
|
図2
(a) グルタミン酸投与でスパインを刺激すると(矢頭)、頭部体積が増大する。(b) スパイン頭部体積変化の時間経過。隣接したスパインは影響を受けない。(c)
小さなスパインでは体積増大が長時間持続する。(d) 大きなスパインでは30分もすると元の大きさに戻る。
|
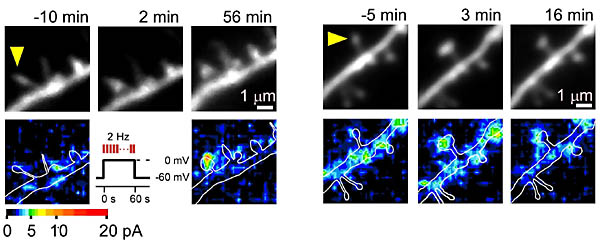
最後に、この刺激によって、形態変化と同時に、LTP、すなわちAMPA受容体反応の増強が起こっているかを調べた。細胞内状態をインタクトに保つため、GFPを発現している錐体細胞に対して、穿孔パッチクランプを行った。1分間、細胞を脱分極(0
mV)させ、その間、2 Hzで狙ったスパインに2光子励起法でグルタミン酸を投与した(図3)。すると、マグネシウム非存在下で行った実験結果と同じように、刺激直後から刺激したスパインのみが大きくなった。このとき、AMPA受容体の電流反応のマッピングを刺激前後で行うと、大きくなったスパインでのみAMPA受容体反応が増強した。さらにスパイン増大が保持されているものにおいては、同じようにAMPA受容体反応の増強も持続していた。また、この増強は小さいスパインでのみ見られ、大きいスパインでは形もAMPA受容体反応もあまり変化がみられなかった。
|
図3
スパインの蛍光像(上)とグルタミン酸感受性(下)。1画像を256分割したずべての点でケイジドグルタミン酸を活性化し、そのときのAMPA受容体電流を穿孔パッチクランプにより記録した。電流の大きさは色表示してある。Pairing刺激したスパイン(矢頭)のみ頭部が大きくなり、グルタミン酸感受性も増強する(2例)。時間は刺激時を0とした時の時間。 |
3.まとめ
まとめると我々の結果は以下のようになる。
1) 長期増強時には、スパイン頭部体積が増大する。AMPA受容体反応の変化と同じように、刺激直後から頭部増大が起こり、持続する。
2) スパインは1個ごとに個別にヘッブの学習法則に従い、記憶素子の最小単位として振る舞いうる。
3) 大きなスパインはAMPA受容体を数多く持ち、なおかつ安定である。
小さなスパインはAMPA受容体数は少ないが、大きくなり易い。
4) スパインの大きさの多様性は、AMPA受容体の発現量(記憶の貯蓄量)に
大きなばらつきがあることを示している。
スパインはその構造がAMPA受容体という機能を決定していながら、非常に柔らかく、数秒レベルで大きく形を変える。これはアクチンを含む細胞骨格系の再編成がすばやく起こり、その後の形態の固定化はキナーゼを介した細胞内伝達系が働くことによると考えられる。生きた動物の脳の中で、このようなスパイン形態変化が学習によって起こり、本当に記憶の物理的基盤となっているのかどうか、今後の課題である。
文献
1) Kasai H, Matsuzaki M, Noguchi J, Yasumatsu N and Nakahara H. Structure-stability-function
relationships of dendritic spines. Trends Neurosci. 26: 360-368, 2003.
2) Harris KM, Jensen FE and Tsao B. Three-dimensional structure of dendritic
spines and synapses in rat hippocampus (CA1) at postnatal day 15 and
adult ages: implications for the maturation of synaptic physiology and
long-term potentiation. J Neurosci. 12: 2685-705, 1992.
3) Yuste R and Bonhoeffer T. Morphological changes in dendritic spines
associated with long-term synaptic plasticity. Annu. Rev. Neurosci.
24: 1071-1089, 2001.
4) Matsuzaki M, Ellis-Davies GCR, Nemoto T, Miyashita Y, Iino M and
Kasai H. Dendritic spine geometry is critical for AMPA receptor expression
in hippocampal CA1 pyramidal neurons. Nature Neurosci. 4: 1086-1092,
2001.
|