|
暉搰媏榊乮杒奀摴戝妛丒戝妛堾堛妛尋媶壢丒摑崌惗棟妛島嵗擣抦峴摦妛暘栰乯
丂侾丏尋媶偺攚宨
丂椉娽帇婡擻傪旛偊偨楈挿椶偱偼栐枌偵帇椡偑嬌傔偰傛偄晹暘偑敪払偟偨丅楈挿椶偑帇妎懳徾暔傪尒偮傔偨応崌丄偦偺搢棫幚憸偼偙偺拞怱鈢偵寢偽傟傞丅拞怱鈢椞堟偼僸僩偱偼帇妏偵偟偰栺1.6亱偱旕忢偵嫹偔丄偦傟偐傜傢偢偐悢搙偢傟傞偩偗偱帇椡偼嬌抂偵掅壓偡傞丅偦偺偨傔俁師尦嬻娫偐傜偺帇妎忣曬傪揔愗偵庢傝崬傓偨傔偵偼丄惓妋側娽媴塣摦傪峴偭偰嬻娫撪偱堄枴偺偁傞庬乆偺帇妎懳徾傪椉娽偺拞怱鈢偵寢憸偝偣丄曔傜偊傜傟偨栐枌憸傪僽儗偢偵曐帩偟側偗傟偽側傜側偄丅帇妎懳徾偑恎嬤偺嬻娫傪備偭偔傝摦偔応崌偺悘堄揑捛愓娽媴塣摦偲偟偰妸摦惈娽媴塣摦(smooth
pursuit)偲鐃鐂塣摦(vergence)偺俀偮偺惂屼宯偑庡偵巊傢傟傞丅妸摦惈娽媴塣摦偼丄巹偨偪偐傜傎傏摍嫍棧偺慜妟柺偱摦偔帇妎懳徾傪丄嵍塃偺娽媴傪摨偠曽岦偵smooth偵摦偐偟偰捛愓偡傞嫟摨惈娽媴塣摦偱偁傞丅偙偺塣摦傪嬱摦偡傞帇妎擖椡偺庡梫惉暘偼丄帇昗偺栐枌忋偺懍搙偲丄偦傟傪捛愓偡傞娽媴懍搙偲偺嵎乮栐枌懍搙岆嵎乯偱偁傞丅鐃鐂塣摦偼墱峴偒曽岦偱埵抲偺堎側傞懳徾暔偐傜偺帇妎忣曬傪庢傝崬傓応崌偵嵍塃偺娽媴傪斀懳曽岦偵摦偐偡塣摦偱偁傞丅偙偺応崌偺帇妎忣曬偼椉娽帇嵎偲屇偽傟丄帇妎懳徾偺椉娽栐枌傊偺搳塭晹埵偲嵍塃娽偺拞怱鈢偲偺嫍棧偵懳墳偟丄栐枌憸偺埵抲忣曬偵側傞丅偙傟傑偱偺尋媶偱偼丄妸摦惈娽媴塣摦偲鐃鐂塣摦偼栐枌偵搳塭偝傟傞帇妎忣曬偺堎側偭偨惉暘傪巊偄丄擼撪偺堎側偭偨椞堟偑屄暿偵張棟偟丄偦傟偧傟偺塣摦巜椷傕擼撪偺堎側偭偨椞堟偱張棟偝傟偰擼姴偺嵟廔弌椡偺抜奒偱椉塣摦巜椷偑摑崌偝傟傞偙偲偑曬崘偝傟偰偒偨侾乯丅偙偺乬屄暿張棟乭偺掕愢偼19悽婭屻敿偺僪僀僣偱偺Helmholtz偲Hering偲偺楌巎揑戝榑憟偵慿傞2乯丅擼偑偳偺傛偆偵偟偰嵍塃娽偺嫤挷塣摦傪惂屼偡傞偐偵偮偄偰Helmholtz偼丄擼偼杮棃丄嵍塃娽傪屄暿偵惂屼偟椉娽偺嫤挷塣摦偼惗屻妉摼偝傟傞傕偺偱偁傞偲偄偆壖愢傪棫偰偨丅偙傟偵懳偟Hering偼丄擼偼惗摼揑偵椉娽偵懳偟偰丄摨堦曽岦乮嫟摨惈娽媴塣摦乯偁傞偄偼斀懳曽岦乮鐃鐂塣摦乯偺摨堦偺巜椷怣崋傪嵍塃娽偵憲傞偲偄偆壖愢傪採彞偟偨丅椺偊偽嵍偺栚慄忋傪帇妎懳徾偑嬤偯偔偲偒嵍娽偼摦偐側偄乮恾侾a)丅偙傟偼鐃鐂塣摦巜椷偲嫟摨塣摦巜椷偑媡曽岦偵側傞偨傔憡嶦偝傟傞寢壥偱偁傞偲愢柧偟偨丅偙偺尒帠側愢柧偵傛傝丄Hering偺壖愢偼Hering偺朄懃(1868)偲偟偰丄偦偺屻丄崱擔傑偱恄宱妛偺嫵壢彂偺掕愢偵側偭偰偄傞1,2乯丅
丂妸摦惈娽媴塣摦偺敪尰偵偼戝擼旂幙偺峀偄椞堟偑娭傢傞侾乯丅栐枌忋偺帇妎懳徾偺懍搙忣曬偼帇妎栰偐傜MT栰(middle
temporal area)偍傛傃MST栰(middle superior temporal area)傪宱丄堦晹偼捈愙偵丄嫶妀乮攚奜懁嫶妀乯傪宱偰彫擼曅梩椞堟傊搳幩偟丄擼姴偺恄宱愊暘夞楬傪宱偰奜娽嬝塣摦僯儏乕儘儞偵弌椡偝傟傞丅懠偼丄慜摢梩娽媴塣摦娭楢椞堟乮慜摢娽栰屻晹椞堟丄曗懌娽栰乯傪宱偰嫶妀乮攚奜懁嫶妀丄嫶旐奧栐條妀乯偵帄傝丄嫶旐奧栐條妀偐傜偼彫擼拵晹丒幒捀妀傪宱偰丄慜掚恄宱妀偵帄偭偰擼姴夞楬偵崌棳偡傞丅鐃鐂塣摦偺宱楬偼枹偩偵妋掕偟偰偄側偄丅妸摦惈娽媴塣摦偺夞楬偺拞偺暋悢偺椞堟偱鐃鐂塣摦墳摎僯儏乕儘儞偑曬崘偝傟偰偄傞偑丄妸摦惈娽媴塣摦墳摎僯儏乕儘儞偲鐃鐂塣摦墳摎僯儏乕儘儞偼堎側傞偙偲偑曬崘偝傟丄慜摢娽栰偱傕偙偺俀庬椶偺娽媴塣摦偺惂屼偵偼堎側傞晹埵偑娭傢傞偙偲偑曬崘偝傟偨1丄3乯丅偨偩偟MST栰偱偼峀偄帇栰偵帇妎僷僞乕儞傪採帵偟偦傟傪丄慜妟柺偱媫懍偵摦偐偟偨応崌偺斀幩惈嫟摨惈娽媴塣摦乮ocular
following丄捛廬娽媴塣摦乯偵墳摎偡傞僯儏乕儘儞偺堦晹偑丄摨條偺帇妎僷僞乕儞傪墱峴偒曽偵摦偐偟偨偲偒偺斀幩惈鐃鐂塣摦偵傕墳摎偡傞偙偲偑曬崘偝傟偨4乯丅
丂擔忢惗妶偱偼丄椺偊偽帺暘偑曕偒側偑傜恎嬤偵曕偄偰偄傞恖偺昞忣傪撉傒偲傟傞傛偆偵妸摦惈娽媴塣摦偼丄帇慄乮嬻娫撪娽媴乯塣摦偲偟偰幚峴偝傟傞丅廬偭偰慜摢娽栰偺栶妱傪棟夝偡傞偨傔偵偼丄慜摢娽栰偑悘堄揑側帇慄塣摦偺嵺偵幚嵺偵偳偺傛偆側怣崋傪帩偮偐傪傑偢柧傜偐偵偟側偗傟偽側傜側偄偲偄偆娤揰偐傜丄杮尋媶幰傜偼摢晹傪屌掕偟偨僯儂儞僒儖傪孭楙偟丄妸摦惈娽媴塣摦壽戣偩偗偱側偔慜掚夞揮巋寖傕梡偄偰娽鈢撪娽媴塣摦偲帇慄塣摦傪槰棧偝偣偰慜摢娽栰屻晹椞堟偺妸摦惈娽媴塣摦僯儏乕儘儞偺墳摎摿惈傪挷傋偰偒偨丅偦偺寢壥丄戝懡悢乮栺俈妱乯偼帇慄塣摦偺懍搙偵墳摎偟丄偦偺嵟揔墳摎曽岦偼妸摦惈娽媴塣摦偺嵟揔曽岦偲堦抳偟丄慡懱偲偟偰偁傜備傞曽岦偑慜摢娽栰屻晹椞堟偱嵞尰偝傟偰偄傞偙偲傪柧傜偐偵偟偨5乯丅傑偨俀屄偺帇昗傪梡偄丄惷巭偝偣偨1屄傪僒儖偵屌帇偝偣丄懠曽傪庬乆偺曽岦偵摦偐偟娽媴塣摦傪婲偙偝偣偢偵帇妎墳摎傪挷傋偨寢壥丄偙傟傜僯儏乕儘儞偺夁敿悢偼栐枌忋偺帇昗偺塣摦曽岦偲懍搙忣曬傪旛偊偰偄傞偙偲傪柧傜偐偵偟偨5乯丅偝傜偵偙傟傜僯儏乕儘儞偼丄梊應揑側妸摦惈娽媴塣摦偲懳墳偟偰墳摎偡傞偩偗偱側偔丄惷巭帇昗偺屌帇拞偵戞2偺帇昗塣摦偵懳偡傞梊應惈偺帇妎墳摎傕帵偡偙偲偑柧傜偐偵側偭偨6乯丅妸摦惈娽媴塣摦偲鐃鐂塣摦偱偼丄恎嬤偺嬻娫撪傪帇妎懳徾偑備偭偔傝摦偔応崌偵丄帇慄塣摦偲梊應傪岠壥揑偵巊偆偲偄偆嫟捠梫慺傪傕偮偨傔丄椉娽媴塣摦宯偺摑崌偑慜摢梩偱婲偙偭偰偄傞壜擻惈傪峫偊偰杮幚尡傪峴偭偨偲偙傠丄埲壓偵帵偡傛偆偵丄慜摢娽栰屻晹椞堟偑俁師尦嬻娫偱偺娽媴塣摦怣崋偺宍惉偵廳梫側栶妱傪壥偨偡偲偄偆怴偟偄寢壥傪摼偨7乯丅
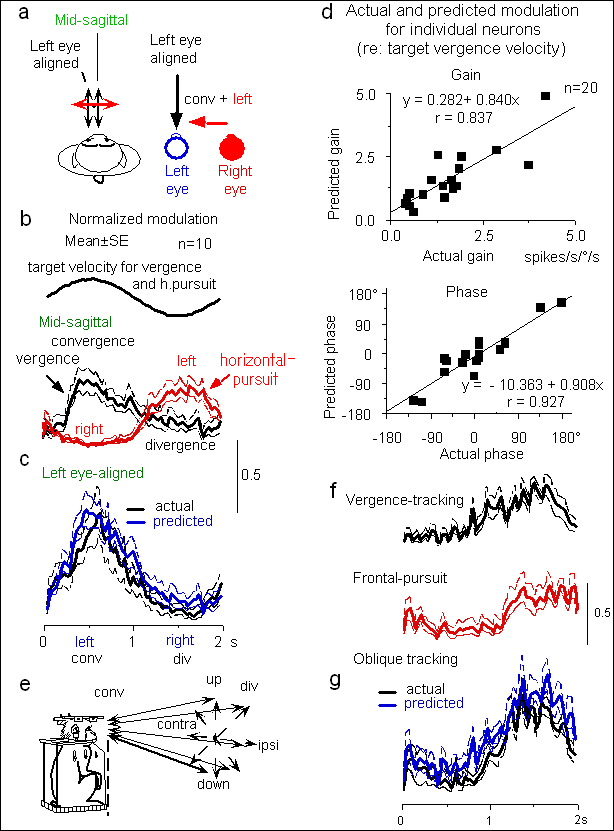 |
| 恾1丏慜摢娽栰屻晹椞堟僯儏乕儘儞偺妸摦惈娽媴塣摦偲鐃鐂塣摦偵懳偡傞墳摎偲椉墳摎偺慄宍壛嶼丅a, e:帇昗掓帵偺柾幃恾丅倐,
c: 鐃鐂塣摦偲悈暯妸摦惈娽媴塣摦偵懳偡傞暯嬒壔偟偨墳摎丅們:嵍栚慄忋偺旕懳徧惈鐃鐂塣摦偵偍偗傞妸摦惈娽媴塣摦偲鐃鐂奐嶶塣摦墳摎偺慄宍壛嶼丅倓丗屄乆偺僯儏乕儘儞偵偍偗傞幚嵺偺墳摎偲慄宍壛嶼偐傜摼傜傟偨梊應抣偲偺斾妑丅倖丗壖憐帇昗傪梡偄偨帪偺墱峴偒曽岦偲慜妟柺偱偺墳摎偲壖憐嬻娫偱偺嵟揔曽岦偺墳摎偲慄宍壛嶼偵傛傞梊應偝傟傞墳摎丅暥專俈乯丅愢柧杮暥丅
|
丂俀丏尋媶偺奣梫
丂摢晹傪屌掕偝傟偨俀摢偺僒儖偺娽慜偵偼悅捈偲悈暯偺俀庬椶偺僗僋儕乕儞偑採帵偝傟丄僗僋儕乕儞忋偺儗乕僓乕僗億僢僩傪栚偱捛愓偡傞傛偆孭楙偝傟偰偄傞丅悅捈僗僋儕乕儞傪梡偄偰慜摢娽栰屻晹椞堟偐傜妸摦惈娽媴塣摦墳摎傪挷傋丄摨堦僯儏乕儘儞傪婰榐偟側偑傜丄僒儖偺旲偺崅偝偵掓帵偟偨悈暯僗僋儕乕儞傪梡偄偰鐃鐂塣摦偵懳偡傞墳摎傪挷傋偨丅悈暯僗僋儕乕儞忋偺僗億僢僩偼丄僒儖偐傜尒偰惓尫攇忬偵墱峴偒曽岦偵摦偐偝傟偨丅傑偨塼徎僔儍僢僞乕晅偒娽嬀傪梡偄偰僐儞僺儏乕僞乕夋柺忋偵俁師尦壖憐嬻娫撪傪摦偔帇昗傪採帵偟丄偦傟傪妸摦惈娽媴塣摦偲鐃鐂塣摦偱捛愓偝偣偨乮恾侾e乯丅
丂慜摢娽栰屻晹椞堟偺122屄偺壽戣娭楢僯儏乕儘儞偵偮偄偰丄妸摦惈娽媴塣摦偲鐃鐂塣摦傪屄暿偵挷傋偨寢壥丄戝懡悢乮66亾乯偼椉娽媴塣摦偵墳摎偟丄25%偼妸摦惈娽媴塣摦偺傒偵丄9%偼鐃鐂塣摦偺傒偵墳摎偟偨丅椉娽媴塣摦偵墳摎偟偨僯儏乕儘儞偺夁敿悢偱敪幩偺僺乕僋偼鐃鐂偁傞偄偼奐嶶塣摦偺嵟戝懍搙偵傎傏堦抳偟偨丅偙傟傜偼慜愡偱弎傋偨傛偆偵妸摦惈娽媴塣摦帪偵偼摿掕偺嵟揔墳摎曽岦傪帩偪丄偙偺曽岦偼懡悢偺僯儏乕儘儞偱慡懱偲偟偰偁傜備傞曽岦偑嵞尰偝傟偰偄偨丅偦偙偱妸摦惈娽媴塣摦偺嵟揔墳摎曽岦偑悈暯偺僯儏乕儘儞孮偵偮偄偰妸摦惈娽媴塣摦偲鐃鐂塣摦偑梫媮偝傟傞帇昗捛愓壽戣拞偺墳摎傪丄屄乆偺娽媴塣摦壽戣偺墳摎偲斾妑偟偨椺偑恾1(a-c)偵帵偟偰偁傞丅偙偺僯儏乕儘儞孮偼椉娽拞墰慄忋偱帇昗偑嬤偯偔偲偒乮鐃鐂乯偵敪幩偑憹偊丄偐偮嵍岦偒偺妸摦惈娽媴塣摦偱敪幩傪憹壛偟偨乮恾1a,b)丅墱峴偒帇昗偑嵍偺栚慄忋偱嬤偯偔偲偒乮恾1a)丄塃栚偼鐃鐂塣摦傪婲偙偟丄偐偮嵍偵摦偔丅偙偺塣摦帪偺幚嵺偺僯儏乕儘儞墳摎偼丄恾1c偵帵偡傛偆偵椉娽拞墰慄忋偱帇昗偑嬤偯偔偲偒偺敪幩偲嵍岦偒妸摦惈娽媴塣摦偱偺敪幩偺慄宍壛嶼偲堦抳偟偨丅屄乆偺僯儏乕儘儞偵偮偄偰妸摦惈娽媴塣摦偲鐃鐂塣摦偑壛嶼偝傟傞曽岦偱偺帇昗巋寖偵懳偡傞墳摎偺埵憡偲棙摼傪丄幚嵺偺墳摎偲梊應偝傟傞墳摎偱斾妑偟偨偺偑恾1d偵傑偲傔偰偁傞丅孹偒偑0.8-0.9丄憡娭學悢0.84-0.93偱椉幰偑傛偔堦抳偡傞偙偲傪帵偡丅偙傟傜偺僯儏乕儘儞偼墱峴偒帇昗偑塃偺栚慄忋偱嬤偯偔応崌偱傕偦傟傜偺墳摎偼椉娽媴塣摦偺壛嶼偱愢柧偱偒偨丅偙偺応崌丄嵍娽偑塃偵摦偔偙偲偵側傞偺偱丄妸摦惈娽媴塣摦墳摎偲鐃鐂塣摦墳摎偼憡嶦偝傟傞偙偲偵側傝丄摿偵妸摦惈娽媴塣摦帪偵敪幩偑巭傑偭偨偑丄嵶朎撪揹埵曄壔傪悇掕偟晧偺僯儏乕儘儞妶摦傪壖掕偡傞偙偲偵傛傝墳摎偑憡嶦偝傟傞曽岦偱傕慄宍壛嶼偱愢柧偱偒偨7乯丅嵟揔曽岦偑悈暯曽岦埲奜偺僯儏乕儘儞偵偍偗傞妸摦惈娽媴塣摦偲鐃鐂塣摦偲偺壛嶼偼丄僐儞僺儏乕僞乕夋柺忋偵壖憐帇昗傪採帵偟丄偦傟傪捛愓偝偣傞偙偲偵傛偭偰挷傋偨乮恾1e乯丅偙偺応崌傕慜妟柺偱偺嵟揔曽岦偲墱峴偒曽岦偱偺墳摎偺嵟揔曽岦偼傎傏堦抳偟丄墳摎偺戝偒偝傕恾1(f,g)偵帵偡傛偆偵椉娽拞墰慄忋偱偺墳摎偲慜妟柺偱偺墳摎偺慄宍壛嶼偱愢柧偱偒偨丅埲忋偺寢壥偼丄妸摦惈娽媴塣摦偲鐃鐂塣摦偺摑崌偑慜摢娽栰偱婲偙偭偰偄傞偙偲丄偝傜偵偦偺椞堟偑3師尦嬻娫偵嵟揔儀僋僩儖傪帩偮娽媴塣摦怣崋傪嵞尰偡傞偙偲傪帵偡丅偙偺嵟揔儀僋僩儖傪柾幃揑偵帵偟偨偺偑恾2乮a,b乯偱偁傞7乯丅
丂偙傟傜偺僯儏乕儘儞婰榐晹埵乮恾2c乯偵GABA嶌摦栻muscimol(10兪g) 傪旝検拲擖偟偰丄偦偺椞堟傪晄妶惈壔偟偨偲偙傠丄妸摦惈娽媴塣摦偺懍搙偑掅壓偟丄棙摼乮娽媴懍搙/帇昗懍搙乯偼傎傏0.5-0.6偵傑偱掅壓偟偨丅偙傟傜偺塣摦忈奞偵敽偄鐃鐂塣摦傕摨條偵忈奞偝傟偨7乯丅
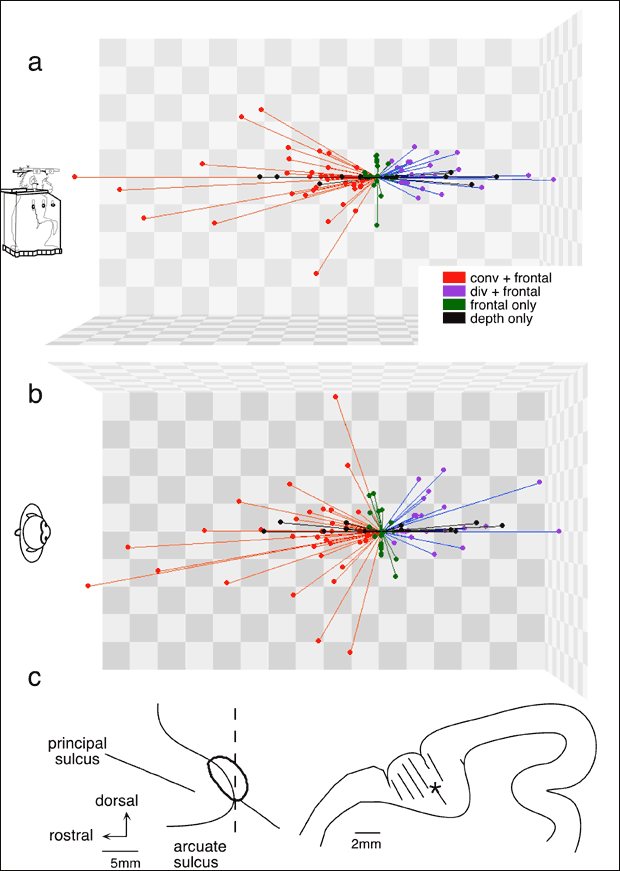 |
| 恾2丏慜摢娽栰屻晹椞堟偵嵞尰偝傟傞俁師尦嬻娫偵偍偗傞娽媴塣摦怣崋丅揰偲捈慄偼屄乆偺僯儏乕儘儞偺嵟揔墳摎儀僋僩儖傪丄婲巒揰傪娽慜偺堦揰偲偟偰帵偡丅a丄懁柺恾丅倐丄忋偐傜尒偨恾丅們丄1摢偺僒儖偺婰榐晹埵丅仏偼muscimol拲擖晹埵丅暥專俈乯丅
|
丂俁丏椉娽帇婡擻偺敪尰偵昁梫側椉娽偺嫤挷塣摦偺惂屼偵偍偗傞慜摢娽栰屻晹椞堟偺栶妱
丂慜摢娽栰屻晹椞堟偵偼smooth側帇慄塣摦傪敪尰偝偣傞偨傔偵昁梫側帇昗塣摦偺栐枌忣曬丄娽媴塣摦忣曬丄慜掚忣曬偑嵞尰偝傟偰偍傝丄偙偺椞堟偺妸摦惈娽媴塣摦僯儏乕儘儞偺戝懡悢偼丄帇慄塣摦偺懍搙忣曬傪旛偊偰偄傞5乯丅偝傜偵偙傟傜慜妟柺偱偺帇慄塣摦怣崋偵壛偊偰墱峴偒曽岦偺娽媴塣摦怣崋傕壛嶼偝傟傞偙偲偵傛傝7乯丄偙偺椞堟偼俁師尦嬻娫偱偺帇昗捛愓傪揔愗偵峴偆偨傔偺椉娽偺嫤挷塣摦偺惂屼偵昁梫偱偁傞偲夝庍偝傟傞丅偙偺夝庍偼丄偙偺椞堟偺晄妶惈壔偑慜妟柺偱偺帇慄塣摦偺傒側傜偢鐃鐂塣摦傕忈奞偡傞偙偲偵傛傝巟帩偝傟傞7乯丅媩忬峚慜暻偺徴摦惈娽媴塣摦僯儏乕儘儞傕傑偨丄椉娽帇嵎忣曬傪傕偮偙偲偑曬崘偝傟偰偄傞偺偱8乯丄慜摢娽栰椞堟偺栶妱偵偮偄偰丄偙傟傑偱偺慜妟柺偱偺娽媴塣摦偺惂屼偱偼側偔丄俁師尦嬻娫偵偍偗傞椉娽偺嫤挷塣摦偺惂屼偲偄偆怴偨側棟夝偑昁梫偱偁傞丅偙傟傑偱偺娽媴塣摦儌僨儖偱偼丄妸摦惈娽媴塣摦偲鐃鐂塣摦偑暿偺僔僗僥儉偱偁傞偲偄偆掕愢偺偨傔丄椉幰傪摑崌偟偨儌僨儖偼慡偔懚嵼偟側偄丅杮寢壥偐傜俁師尦嬻娫偺帇昗塣摦偍傛傃偦傟傪捛愓偡傞娽媴塣摦偵偮偄偰丄慜妟柺偲墱峴偒曽岦傪摑堦揑偵愢柧弌棃傞怴偟偄嬻娫儌僨儖偺奐敪偑昁梫偵側傞丅
丂惓妋側娽媴塣摦偵傛傞帇妎忣曬偺揔愗側庢傝崬傒偼丄3師尦嬻娫偱偺庬乆偺塣摦峴摦丄摿偵恎嬤偺暔懱偵懳偡傞摢晹傗庤巜摍偺塣摦偵昁恵偱偁傞丅慜摢娽栰偺俁師尦嬻娫偵懳偡傞娽媴塣摦怣崋偑丄庤傪娷傔偨懱偺塣摦惂屼偵傕巊傢傟傞壜擻惈偑帵嵈偝傟傞9丄10乯丅
暥專
1)Leigh R et al. The neurology of eye movements. 3rd
ed. Oxford University Press New York, 1999
2)King WM et al. New Anat (2000) 261: 153-161
3)Gamlin PD et al. Nature (2000) 407: 1003-1007
4)Takemura A et al. J Neurophysiol (2001) 85: 2245-2266
5)Fukushima K et al. J Neurophysiol (2000) 83: 563-587
6)Fukushima K et al. Exp Brain Res (2002) 145: 104-120
7)Fukushima K et al. Nature (2002) 419: 157-162,
8)Ferraina S et al. J Neurophysiol (2000) 83: 625-629
9)Kakei S et al. Nat Neurosci (2001) 4:1020-1025
10)Mushiake H et al. J Neurophysiol (1997) 78:567-571
|
