| 根岸 学(京都大学・大学院生命科学研究科・生体システム学分野)
学習や記憶などの複雑な脳機能を可能にしているのは、神経細胞が神経突起をのばし、互いに接着することにより形成される複雑な神経回路の存在である。神経細胞は特異な極性を持つ細胞であり、通常、細胞体から1本の軸索と複数の樹状突起を伸長する。この特異な細胞形態が神経回路形成による高次な脳機能発現の基盤となっている。近年、この神経突起形成・伸長の調節にRhoファミリー低分子量G蛋白質が重要な役割を果たしていることが明らかとなってきた1)。そして、精神遅滞などの遺伝性神経疾患の原因遺伝子としてRhoファミリーの情報伝達系に関わる分子が次々と同定されてきたことから、神経回路形成におけるRhoファミリーの生理的役割の意義がクローズアップされている。
Rhoファミリーにはたくさんの種類があり、現在までにほ乳類で少なくとも14種類が見つかっている。一般的にRhoファミリーは他の低分子量G蛋白質と同様に、グアニンヌクレオチド交換因子(GEF)によりGTP結合型となり活性化され、下流の特異的なエフェクターに結合し、様々な機能を発揮し、GAPによりGTP水解が促進され、不活性型となる。Rhoファミリーの中では、RhoA、Rac1、Cdc42で最もよく研究が進んでおり、Rac1とCdc42は神経突起伸長に、RhoAは突起の退縮に関与することがよく知られている。また、それぞれの下流のエフェクター分子も多数同定され、Cdc42はN-WASPを、Rac1はPAKを介して突起の伸長を促進すること、RhoAはRhoキナーゼを介して突起の退縮や成長円錐の消失を引き起こすことが分かってきた。一方、Rho、Rac、Cdc42の様々なGEFが見いだされているが、RacとCdc42に関してどのような機構でその活性が制御されているのかはあまりよく分かっていない。
神経突起形成の研究において、PC12細胞はよく研究されてきたモデル系で、NGFが細胞膜上のチロシンキナーゼ型受容体、TrkAに結合し、Rasを活性化して神経突起を伸長する。NGFによる神経突起伸長にRac1とCdc42の活性が必須であることは、Rac1とCdc42のドミナントネガティヴ体がNGFによる突起伸長を抑制することから推察されていた。しかし、活性型Rac1やCdc42を発現させてもラッフリングやフィロポディアが細胞膜全体で形成されるだけで神経突起は形成されず、Rac1やCdc42の上流で何らかの活性の制御機構の存在が示唆された。以前、我々はRhoファミリーの中でRhoGという別のG蛋白質がNGF/Rasの下流でRac1とCdc42を活性化し、神経突起を伸長させることを見いだした2)。Rac1とCdc42はそのエフェクター分子で直接細胞骨格系を制御し、神経突起を伸長させるが、RhoGはこれらのG蛋白質の上位の制御分子であり、NGF/Rasのシグナルを受けて突起形成部位でRac1とCdc42のGTPaseサイクルを促進することによりダイナミックな神経突起伸長を推進するものと推定された。RhoGの活性化経路として、最近、DubantのグループがRhoGのGEF、TrioがNGF/Rasの下流でRhoGを活性化し、神経突起を伸長することを報告した3)。しかし、RhoGによるRac1やCdc42の活性化経路はミッシングリンクであった。
そこで、我々は、この活性化機構を明らかにすべく、Rac1やCdc42を活性化するRhoGのエフェクターを見いだすため、酵母のtwo-hybrid法を用いてラット脳のcDNAライブラリーをスクリーニングし、Elmoという分子を同定した。ElmoはRac特異的なGEF、Dock180に結合する分子として見いだされた分子であり、ファゴサイトーシスや細胞移動に必須の分子であることが明らかにされていた4,5,6)。Dock180は最初、MatsudaらによりCrkに結合する分子として同定され、Racに直接結合し活性化することが示された7,8)。通常、RhoファミリーのGEFにはその触媒部位であるDH領域とPH領域がタンデムに存在するが、Dock180はDH領域を持たず、代わりにDHR2という全く新しい触媒領域を持つ分子である。一方、ElmoはC末端側にPH領域を持つ以外、既知の領域は見いだされていない。ElmoはそのPH領域を含むC末端側でDock180のN末端にあるSH3領域と結合し、安定な複合体を形成し、細胞質に存在する。ElmoはRhoGの特異的なエフェクターであり、他のRhoファミリー、RhoA、Rac1、Cdc42などは全く結合しない。活性型RhoGはElmoのN末端側に結合し、Elmo/Dock180複合体を細胞膜に移行させ、RhoG/Elmo/Dock180三者複合体を形成し、細胞膜上でRac1を活性化することが分かった。ElmoのC末端のDock180結合領域を欠失した変異体、N末端のRhoG結合部位を欠失した変異体、Dock180の触媒活性を欠失した変異体は全てNGF及びRhoGによる神経突起伸長を抑制したことから、PC12細胞において、NGFはRhoG/Elmo/Dock180の新しい経路を介してRac1を活性化し、神経突起を伸長させることが明らかとなった。
一方、NGF及びRhoGはRac1と共にCdc42を活性化する。しかし、Dock180はRac特異的なGEFであり、Cdc42を活性化しない。Dock180のようにDH領域を持たないGEFはヒトで少なくとも11種類見いだされており、CDMファミリーと呼ばれ全て共通の触媒部位、DHR2をもつ。CDMファミリーは4つのサブファミリーに分かれ、その中でZizimin1(DOCK9)はCdc42を活性化するGEFであることが報告された9)。現在、RhoGによるCdc42の活性化にZiziminが関与するのか、また、そのためのRhoGのエフェクターが何かは全く不明であるが、CDMファミリーの分子がRhoG/Cdc42の経路に関わる可能性は十分にあると思う。Elmo/Dock180はC.
elegansからほ乳類まで種を超えて幅広くRacの上流の活性化経路として確認されており、C. elegansではそれぞれCED12/CED5、DrosophilaではDCED12/Mbcと呼ばれている。RhoGにはC.
elegansでMIG-2が対応し、細胞の移動や軸索ガイダンスに必須であることが示された10)。また、DrosophilaではMtlが対応する。このように、RhoG/Elmo/Dock180はRacの活性化経路として種を超えて進化の過程で保存されている経路であることがわかる。現在、RhoGのGEFとしてTrio
、Kalirin、Vavの3種類が知られている。その中で、交感神経においてKalirinがRhoGを活性化し、軸索伸長を引き起こすことが示された11)。また、C.
elegansにおいてTrioのホモログであるUNC-73が成長円錐の伸展に必須であることも知られている。これらのことから、Trio/Karilin-RhoG-Elmo-Dock180-Racの経路が神経突起の伸長に極めて重要な役割を果たしていることが示唆される。RhoファミリーG蛋白質は、そのGTPaseサイクルにより神経細胞内で空間的及び時間的に情報のオン・オフを厳密に制御するバイオタイマーとして働いており、その活性制御は神経回路形成において重要なシステムと考えられる。
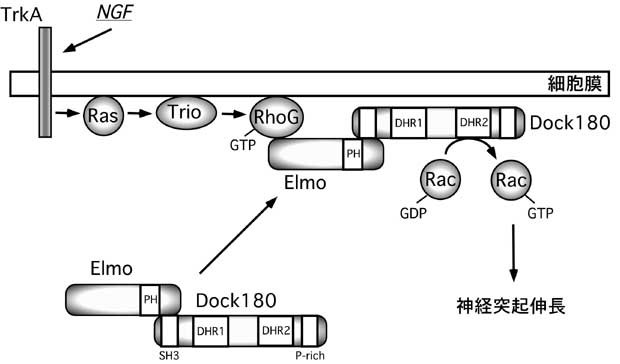 |
図1. NGFによる神経突起伸長における細胞内情報伝達経路
TrioはRhoGのGEFであり、RhoGを活性化し、活性型GTP-RhoGはElmo/Dock180複合体を細胞膜に移行させ、三者複合体を形成し、Rac1を活性化し、神経突起を伸長させる。 |
文献
1) Negishi M, Katoh H. Rho family GTPases as key regulators for neuronal
network formation. J Biochem 132: 157-166, 2002
2) Katoh H, Yasui H, Yamaguchi Y, Aoki J, Fujita H, Mori K, Negishi
M. Small GTPase RhoG is a key regulator for neurite outgrowth in PC12
cells. Mol Cell Biol 20; 7378-7387, 2000
3) Estrach S, Schmidt S, Diriong S, Penna A, Blangy A, Fort P, Debant
A. The human Rho-GEF Trio and its target GTPase RhoG are involved in
the NGF pathway, leading to neurite outgrowth. Curr Biol 12: 307-312,
2002
4) Wu Y-C, Tsai M-C, Cheng L-C, Chou C-J, Weng N-Y. C. elegans CED-12
acts in the conserved CrkII/DOCK180/Rac pathway to control cell migration
and cell corpse engulfment. Dev Cell 1: 491-502, 2001
5) Zhou Z, Caron E, Hartwieg E, Hall A, Horvitz HR. The C. elegans PH
domain protein CED-12 regulates cytoskeletal reorganization via a Rho/Rac
GTPase signaling pathway. Dev Cell 1: 477-489, 2001
6) Gumienny TL, Brugnera E, Tosello-Trampont AC, Kinchen JM et al. CED-12/ELMO,
a novel member of the CrkII/Dock180/Rac pathway, is required for phagocytosis
and cell migration. Cell 107: 27-41, 2001
7) Kiyokawa E, Hashimoto Y, Kobayashi S, Sugimura H, Kurata T, Matsuda
M. Activation of Rac1 by a Crk SH3-binding protein, DOCK180. Genes Dev
12: 3331-3336, 1998
8) Brugnera E, Haney L, Grimsley C, Lu M et al. Unconventional Rac-GEF
activity is mediated through the Dock180-ELMO complex. Nature Cell Biol
4: 574-582, 2002
9) Meller N, Irani-Tehrani M, Kiosses WB, Pozo MAD, Schwartz MA. Zizimin1,
a novel Cdc42 activator, reveals a new GEF domain for Rho proteins.
Nature Cell Biol 4: 639-647, 2002
10) Zipkin ID, Kindt RM, Kenyon CJ. Role of a new Rho family member
in cell migration and axon guidance in C. elegans. Cell 90: 883-894,
1997.
11) May V, Schiller MR, Eipper BA, Mains RE. Kalirin Dbl-homology guanine
nucleotide exchange factor 1 domain initiates new axon outgrowths via
RhoG-mediated mechanisms. J Neurosci 22: 6980-6990, 2002
|
