| 石原健(国立遺伝学研究所・構造遺伝学研究センター)
研究の背景
動物は、外界から様々な刺激を感覚神経で受容している。そのような感覚情報は、中枢神経系で、取捨選択され、記憶などとの照合やそれによる修飾を受けるなど、適切に処理され、最終的には、環境に適応した行動として出力される。このような情報処理システムは、単純な神経系を持つ動物においても、個体が生存していく上で必要不可欠なものである。私達は、このような感覚情報処理に関わる分子メカニズムを明らかにすることを目的として、線虫Caenorhabditis
elegans(C. elegans )を用いた行動遺伝学的解析を進めている。
線虫C.elegans は、S. Brennerによって発生と行動の遺伝学的解析のためのモデル生物として導入された1)。その後、Brennerの弟子達によって、細胞系譜の決定2)、電子顕微鏡による神経回路の全構造の決定3)、全ゲノム配列の決定4)などが進められ、現在ではもっとも多くのことが記述されているモデル生物の1つである。C.
elegans 雌雄同体の成虫の体細胞959個のうち、神経細胞は302個118種類であって2)、それらは約5000個の化学シナプス、600個のギャップジャンクションを介して3)、神経ネットワークを形成している。また、約100MBのゲノム中には約2万個の遺伝子が同定されており4)、これはヒトの約2/3の遺伝子数に相当する。このように、C.
elegans は、遺伝子レベルでの複雑さは保ちながら、解剖学的な複雑さを回避できるモデル生物として、分子、神経回路、行動のそれぞれを結び付けて神経系を解析するのに適している。
これまで、C. elegans において1つの刺激に対する応答に異常がある行動変異体の解析が精力的に行われてきた。例えば、Bargmann(UCSF)らによる誘引性匂い物質に対する変異体の解析によって、感覚神経AWA、AWCの分化メカニズムや感覚受容のシグナル系が明らかになってきた5)、6)。しかし、介在神経などからなる神経回路における情報処理システムに異常がある変異体は見つからず、その分子メカニズムはあきらかではない。そこで、我々は、異なる感覚神経で受容される2つの感覚刺激を同時に与えた際の応答を行動として調べることにより、神経回路における情報処理のメカニズムを明らかにすることを目指した。
研究の概要7)
行動測定系とhen-1 変異体の表現型
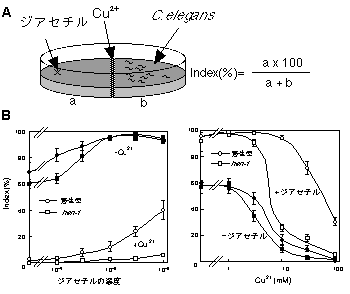
感覚刺激としては、銅イオンと誘引性匂い物質ジアセチルを用いた。C. elegans は、銅イオンを忌避するのに対し、ジアセチルには化学走性を示し寄っていく8)、9)。我々が開発した測定系では、ジアセチルに寄っていくためには、銅イオンのバリアーを越えなければならない(図1A)。この測定系において、C.
elegans は、ジアセチルと銅イオンのそれぞれの濃度に応じて、どちらの刺激に対する応答が優先されるか(銅イオンのバリアーを越えられるか)が異なる。これらの感覚刺激は異なった感覚神経で受容されることから、ジアセチルの感覚情報と銅イオンの感覚情報は、神経回路においてお互いに抑制し合う仕組みがあることを意味している。
|
図1. 感覚情報の統合の測定系と神経回路
A. 感覚情報の統合の測定系。餌のない寒天培地の中央に銅イオンをぬり、片側にジアセチル、片側にC. elegans
の成虫約50匹をおき、1.5時間後の虫の分布を数える。Indexは、すべての虫のうち、銅イオンを越えた虫のパーセンテージとして定義した。
B. hen-1 の感覚情報の統合における異常。左図は、銅イオンの存在下・非存在下でのジアセチルに対する濃度依存性。右図は、ジアセチルの存在下・非存在下での銅イオンに対する濃度依存性。hen-1
変異体は、銅イオンまたはジアセチルの片方のみで刺激した場合の応答は、野生型と変わらないが、両者の存在下での行動は、野生型と大きく異なる。
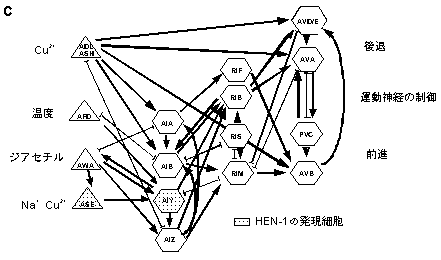
C. 感覚情報処理に関わる神経回路。感覚神経(三角形)から、介在神経(六角型)に化学シナプス(矢印)が形成されている。最も右の列の介在神経は、運動神経を直接制御するコマンドインターニューロンであり、この10数対からなる神経細胞で形成される神経回路で情報が処理されていると推定される。このうち、HEN-1が発現しているAIY神経は感覚情報の統合に必須である。 |
このような感覚情報の統合の測定系において異常を示す変異体hen-1 を同定した。hen-1 変異体では、銅イオンからの忌避応答がジアセチルへの化学走性より優先されるようにみえる。このような表現型を示す原因としては、感覚受容に異常がある可能性があるが、銅イオンからの忌避応答やジアセチルに対する化学走性の応答そのものには異常がなかった(図1B)。従って、感覚受容ではなく、神経回路での情報処理、すなわちジアセチルの感覚情報と銅イオンの感覚情報とがお互いに抑制し合う仕組みに異常がある可能性が考えられた。次に、このほかの高次行動を解析したところ、一種の連合学習に異常があることが分かった。C.
elegans は、NaClに化学走性を示すが、餌がない条件下でNaClにさらすと、NaClに対する応答が変化してNaClを忌避するようになる10).。hen-1
変異体においては、この行動の可塑性に異常があり、NaClを忌避するようにはならない。また、C. elegans は、餌の十分ある良い条件で飼育された場合、その飼育温度を好み寄っていく温度走性を示す。しかし、同じ飼育温度で、飢餓を経験した場合、その飼育温度を忌避するように行動が変化する(毛利・森私信)11)、12)。hen-1
変異体では、このような飢餓による温度走性行動の可塑性を示さない。これらの2つの学習パラダイムにおいては、飢餓を刺激として用いているが、hen-1
変異体は、飢餓そのものに対する応答には異常がなかったので、連合学習そのものに異常があると考えられる。
このように、hen-1 変異体は、さまざまな刺激の感覚受容そのものには異常がないが、感覚情報の統合と連合学習に異常を示す。感覚情報の統合と連合学習は、一見異なった行動にみえるが、連合学習における記憶の形成は、2つの感覚情報から1つの記憶を形成する過程と考えることもでき、2つの感覚情報から1つの出力という感覚情報の統合と良く似た過程であると考えることができる。
HEN-1タンパク質の働くメカニズム
感覚情報処理に関わる分子機能を明らかにするため、hen-1 変異体の原因遺伝子のポジショナルクローニングを行ったところ、hen-1
遺伝子はLDLaモチーフを1つ持つ分子量約1万の分泌タンパク質をコードしていた。このHEN-1タンパク質の発現を免疫染色により解析したところ、感覚神経ASEと介在神経AIYという2対の神経細胞でのみ発現がみられた(図1C)。また、細胞内での局在を共焦点顕微鏡で観察したところ、神経軸索中、とくに神経結合が多く形成されている神経環と呼ばれる領域で、HEN-1は点状に局在していた。シナプス小胞が輸送されないunc-104
(KIF1Aキネシン相同分子をコードする)変異体では、神経軸索への輸送が観察されなかったことから、シナプス小胞と類似したメカニズムで輸送されていることが推定された。
HEN-1の働く分子機構を解析するため、hen-1 変異体において、様々なプロモーターの制御下で野生型hen-1
遺伝子を発現させ、表現型の回復を調べた。HEN-1が本来発現しているASEやAIY神経で発現させた場合のみならず、ほかの神経でHEN-1を発現させた場合にも、hen-1
変異体の行動表現型を回復させることができたが、体壁の筋肉細胞で発現させた場合には表現型の回復は見られなかった。このことは、HEN-1分子が神経系において分泌されて細胞非自律的に機能していることを示している。次に、熱ショックプロモーターを用いて、発生過程の様々な時期に野生型hen-1
遺伝子を発現させ、hen-1 変異体の成虫における行動表現型の回復を調べた。神経回路が形成される胚発生後期や幼虫期にHEN-1を発現させても表現型の回復はほとんど見られなかったが、行動する直前の成虫期に発現を誘導すると表現型が回復したことから、HEN-1分子は神経回路の形成に働いているのではなく、成熟した神経系において神経機能を制御する分子であると推定された。
HEN-1が発現しているAIY神経は、多くの感覚神経が神経結合を形成している一次介在神経であり、感覚情報処理に関わっていることが予想された(図1C)。そこで、AIY神経の分化に異常があるttx-3
変異体(13)の行動表現型を解析した。ttx-3 変異体は、銅イオンとジアセチルを用いた感覚情報の統合の測定系で、hen-1
変異体と同様の表現型を示した。このことは、AIY神経が、感覚情報の統合に必要であることを示している。
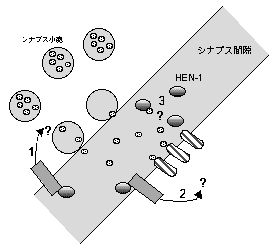
これらのことから、HEN-1分子は、シナプス間隙に分泌され、周りの神経細胞に発現する受容体を介してその神経活動を制御する、あるいは神経伝達物質と相互作用することによりその機能を修飾しているなどにより、神経回路上での情報処理を制御していると考えられた(図2)。
|
図2. HEN-1の働くメカニズム
HEN-1はシナプス間隙に分泌され、周りの神経細胞にある受容体を介して神経機能を修飾している可能性がある。また、神経伝達物質と結合することにより、その機能を修飾している可能性がある。 |
今後の展望
我々は、C. elegans を用いた行動遺伝学により、神経回路における情報処理の制御を担う分子HEN-1を同定した。HEN-1の関わる情報処理の分子機構を明らかにするためには、受容体などHEN-1と相互作用する分子の同定や解析を進めていく必要がある。ヒトやマウスなどでは、HEN-1そのものの相同分子は見つからないが、受容体が同定できれば、その相同分子は同定できる可能性があると考えている。本研究は、C.
elegans が感覚受容の分子メカニズムを解析するのに適しているというだけではなく、比較的単純な情報処理の分子機構の解析にも適したモデル生物であることを示している。今後、C.
elegans の高次行動の遺伝学的解析による新規分子の同定とその機能解析をすすめるとともに、神経活動のイメージングなどとも組み合わせて、神経回路における感覚情報処理の詳細を明らかにしていきたいと考えている。現在、C.
elegans における高次行動の遺伝学は端緒についたところであるが、日本のC. elegans 研究者により、多くの興味深い現象が見出されてきており、今後新たな分子メカニズムが明らかになることが期待される。このようにしてC.
elegans において得られた知見は、ヒトなどの高等生物の複雑な神経回路における情報処理を解析するための重要な基盤となるものと考えている。
本研究は、飯野雄一先生(東京大学遺伝子実験施設)、毛利亮子先生,森郁恵先生(名古屋大学理学部)、安藤恵子先生,三谷昌平先生(東京女子医科大学)、桂勲先生(国立遺伝学研究所)との共同研究である。この場を借りて感謝いたします。図の一部は実験医学編集部のご好意で転載いたしました。
文献
1) Brenner S. : Genetics, 77: 71-94, 1974
2) Sulston J.E. & Horvitz H.R. : Dev Biol, 56: 110-156, 1977
3) White J.G. et al : Philosophical Transactions of the Royal Society
of London, 314B: 1-340, 1986
4) The C. elegans Sequencing Consortium : Science 282:2012-8.
5) Bargmann C.I. : Science, 282: 2028-2033, 1998
6) L'Etoile N.D. & Bargmann C.I. : Neuron, 25: 575-586, 2000
7) Ishihara T. et al : Cell, 109: 639-649, 2002
8) Bargmann C.I. et al : Cell, 74: 515-527, 1993
9) Sambongi Y. et al : Neuroreport, 10: 753-757, 1999
10) Saeki S. et al : J Exp Biol, 204: 1757-1764, 2001
11) Mori I. : Annu Rev Genet, 33: 399-422, 1999
12) Hedgecock E M et al. PNAS, 72: 4061-4065 (1975)
13) Hobert O. et al : Neuron, 19: 345-357, 1997
|