崻娸丂妛丂乮嫗搒戝妛丒戝妛堾惗柦壢妛尋媶壢丒惗懱僔僗僥儉妛暘栰乯
丂侾丄 尋媶偺攚宨
丂妛廗傗婰壇側偳偺暋嶨側擼婡擻傪壜擻偵偟偰偄傞偺偼丄恄宱嵶朎偑恄宱撍婲傪怢偽偟丄屳偄偵愙拝偡傞偙偲偵傛傝宍惉偝傟傞暋嶨側恄宱夞楬偺懚嵼偵傛傞丅捠忢丄恄宱嵶朎偼嵶朎懱偐傜侾杮偺幉嶕偲暋悢偺庽忬撍婲傪怢挿偡傞丅幉嶕偼條乆側幉嶕僈僀僟儞僗暘巕偵摫偐傟偰怢挿偟丄栚揑偺僞乕僎僢僩嵶朎偵摓払偟丄僔僫僾僗傪宍惉偟丄暋嶨側恄宱夞楬傪宍惉偡傞丅偙偺恄宱夞楬栐偺婎杮峔憿偼偐側傝惓妋偱尩枾偵偱偒偰偍傝丄偦傟傪壜擻偵偟偰偄傞偺偼幉嶕僈僀僟儞僗暘巕偺桿摫嶌梡偱偁傞丅尰嵼傑偱偵條乆側幉嶕僈僀僟儞僗暘巕偑敪尒偝傟偰偍傝丄偦偺僈僀僟儞僗嶌梡偐傜戝偒偔暘偗偰俀偮偺僌儖乕僾偵暘偗傜傟傞丅侾偮偼幉嶕偵懳偟斀敪嶌梡傪帵偡暘巕偱丄semaphorin乮Sema乯僼傽儈儕乕丄ephrin僼傽儈儕乕傗Slit側偳偑偁傞丅懠曽偼桿堷嶌梡傪帵偡暘巕偱丄netrin傗NGL-1側偳偑抦傜傟偰偄傞丅幉嶕僈僀僟儞僗暘巕偼丄恄宱嵶朎枌忋偵偁傞偦傟偧傟偵摿堎揑側庴梕懱偵寢崌偟丄嵶朎撪偵僔僌僫儖傪揱払偡傞丅尰嵼傑偱偵條乆側僈僀僟儞僗暘巕偺庴梕懱偑僋儘乕僯儞僌偝傟丄偦偺暘巕峔憿偑柧傜偐偵偝傟偰偒偨偑丄偳偺傛偆側婡峔偱幉嶕僈僀僟儞僗嶌梡傪敪婗偡傞偺偐丄偦偺暘巕婡峔偼傑偩晄柧側揰偑懡偄丅
丂Sema僼傽儈儕乕偼慄拵偐傜僸僩傑偱暆峀偔庬傪挻偊偰懚嵼偡傞斀敪嶌梡傪帵偡僈僀僟儞僗暘巕偱偁傞侾)丅Sema偼尰嵼傑偱偵俈偮偺僋儔僗偵暘椶偝傟偰偍傝丄偦偺拞偱摿偵Sema3偲Sema4偺僋儔僗偑嵟傕傛偔尋媶偝傟偰偄傞丅僋儔僗偵傛傝丄暘斿宆偲嵶朎枌寢崌宆偑偁傝丄Sema3偼暘斿宆偱丄Sema4偼嵶朎枌侾夞娧捠宆偱偁傞丅偟偐偟丄Sema4D偼僾儘僥傾乕僛偵傛傝愗抐偝傟偰暘斿宆偵側傞偲偄偆曬崘偑偁傞丅崱傑偱偵丄Sema偺庴梕懱偺偄偔偮偐偑摨掕偝傟偰偍傝丄係偮偺僒僽僌儖乕僾丄Plexin-A丄B丄C丄D偵暘偐傟傞丅Sema3偼neuropilin偲偄偆嵶朎撪椞堟偑嬌傔偰抁偄暘巕偵寢崌偟丄neuropilin偑Plexin-A偲夛崌偟丄Plexin-A偺嵶朎撪椞堟傪夘偟偰僔僌僫儖揱払傪峴偆丅堦曽丄Sema4D偼Plexin-B1偵捈愙寢崌偟丄嵶朎撪偵忣曬傪揱払偡傞丅Plexin僼傽儈儕乕偺拞偱丄偦偺嵶朎撪忣曬揱払婡峔偺尋媶偑嵟傕恑傫偩偺偼丄Plexin-B偱偁傞丅Plexin-B偺C枛抂偵偼丄PDZ寢崌儌僠乕僼偑偁傝丄嵟嬤丄掅暘巕検G抈敀幙偺RhoA偺妶惈壔場巕乮GEF乯偺侾偮丄PDZ-RhoGEF偑Plexin-B偺C枛抂偵寢崌偟丄RhoA傪妶惈壔偟偰戅弅嶌梡傪敪婗偡傞偙偲偑帵偝傟偨2)丅偟偐偟丄Plexin-B偺C枛抂偺PDZ寢崌儌僠乕僼偼懠偺僒僽僌儖乕僾Plexin-A丄C丄D偵偼懚嵼偣偢丄傑偨丄Plexin-B偱傕傎擕椶偵偼偁傞偑丄僔儑僂僕儑僂僶僄傗慄拵偺Plexin偵偼側偔丄PDZ-RhoGEF偵傛傞RhoA妶惈壔偼丄Plexin僼傽儈儕乕慡懱偺斀敪嶌梡偺晛曊揑側暘巕婡峔偲偼峫偊傜傟側偐偭偨丅Plexin僼傽儈儕乕偺嵶朎撪椞堟偵偼庬傪挻偊偰傛偔曐懚偝傟偰偄傞俀偮偺椞堟丄C1偲C2偑偁傞丅偙偺攝楍偼丄掅暘巕検G抈敀幙偺妶惈梷惂暘巕偱偁傞GAP乮GTPase
activating protein乯偲掅偄側偑傜憡摨惈偑偁傞偙偲偑埲慜偐傜抦傜傟偰偄偨丅偟偐偟丄偙偺椞堟偑偳偺傛偆偵婡擻敪尰偵娭傢傞偐偼慡偔晄柧偱偁偭偨丅
丂俀丄尋媶偺奣梫
丂Rho僼傽儈儕乕G抈敀幙偼嵶朎崪奿偺廳梫側挷愡暘巕偱偁傝丄條乆側幉嶕僈僀僟儞僗暘巕偺婡擻敪尰偵怺偔娭傢偭偰偄傞偙偲偑帵偝傟偰偒偨俁)丅変乆偼丄Rho僼傽儈儕乕G抈敀幙偺拞偱丄擼丒恄宱宯偵摿堎揑偵敪尰偟偰偄傞Rnd1偺恄宱婡擻傪柧傜偐偵偡傞偨傔丄Rnd1偵寢崌偡傞暘巕傪丄峺曣偺two-hybrid朄傪梡偄偰僗僋儕乕僯儞僌偟丄Plexin-B偺嵶朎撪椞堟偵寢崌偡傞偙偲傪尒偄偩偟偨4)丅傑偨丄懠偺僌儖乕僾偑Rnd1偑Plexin-A偵傕寢崌偡傞偙偲傪帵偟偨偙偲偐傜丄Rnd1偑Plexin僼傽儈儕乕偵暆峀偔寢崌偡傞偙偲偑悇嶡偝傟偨丅Rnd1偼懠偺Rho僼傽儈儕乕G抈敀幙偲堎側傝丄忢帪妶惈宆偱丄埨掕偵忢偵Plexin偵寢崌偟偰偄傞丅Rnd1偺拞悤恄宱宯偱偺敪尰傪挷傋傞偲丄戀惗俀侽擔崰偐傜敪尰偟巒傔丄惗屻侾係擔偐傜俀侽擔崰崅偄敪尰偑傒傜傟丄偦偺屻尭彮偡傞俆)丅敪尰偼擼偺峀偄斖埻偱尒傜傟丄摿偵丄戝擼旂幙傗奀攏偺悕懱嵶朎傗丄彫擼偺僾儖僉儞僄嵶朎偵嬌傔偰嫮偔敪尰偟偰偄偨丅偙偺偙偲偐傜丄Rnd1偑恄宱夞楬宍惉婜偵偙傟傜偺恄宱嵶朎偱壗傜偐偺栶妱傪壥偨偟偰偄傞偙偲偑悇嶡偝傟偨丅Plexin-B1偺嵶朎撪椞堟偱丄Rnd1偺寢崌晹埵偼丄C1偲C2偵嫴傑傟偨椞堟偵偁傝丄偦偺椞堟撪偵偁傞Rac傗Cdc42偺寢崌晹埵偱偁傞CRIB攝楍偵帡偨CRIB條僪儊僀儞偵曄堎傪擖傟傞偲丄Rnd1偼寢崌弌棃側偔側傞丅偲偙傠偱丄C1偲C2偼GAP條偺攝楍傪帩偮偑丄嬤擭丄掅暘巕検G抈敀幙偺條乆側GAP偺寢徎峔憿夝愅偑峴傢傟丄G抈敀幙偲偺寢崌條幃偑柧傜偐偵偝傟偰偒偨俇)丅偦偺寢壥丄GAP偱嫟捠偵傛偔曐懚偝傟偰偄傞丄Arg巆婎傪拞墰偵娷傓俈巆婎偱峔惉偝傟偨Arg儌僠乕僼偑俀偮懚嵼偟丄偙偺primary
Arg儌僠乕僼偲secondary Arg儌僠乕僼偑僽儕僢僕傪宍惉偟偰G抈敀幙偵寢崌偟丄GTPase妶惈傪懀恑偡傞偙偲偑柧傜偐偵偝傟偨丅偦偙偱丄Plexin-B1偺嵶朎撪椞堟偺攝楍傪徻嵶偵夝愅偡傞偲丄primary
Arg儌僠乕僼條偺攝楍偑C1偵丄secondary Arg儌僠乕僼條攝楍偑C2偵懚嵼偡傞偙偲偑傢偐偭偨丅偙偺偙偲偼丄Plexin-B1偺嵶朎撪椞堟偑幚嵺偵掅暘巕検G抈敀幙偺GAP偲偟偰婡擻偡傞壜擻惈傪帵嵈偡傞丅偝傜偵丄C1偲C2偺娫偵Rnd1偺寢崌晹埵偑偁傞偙偲偐傜丄Rnd1偺寢崌偑丄偙偺GAP妶惈敪尰偺挷愡偵寛掕揑側栶妱傪壥偨偟偰偄傞壜擻惈偑峫偊傜傟偨丅師偵丄Plexin-B1偑偳偺掅暘巕検G抈敀幙偺GAP偱偁傞偐偱偁傞偑丄Plexin-B1偑斀敪嶌梡傪帵偡偙偲偐傜丄R-Ras偵懳偡傞GAP偱偼側偄偐偲悇掕偟偨丅偦傟偼丄R-Ras偑僀儞僥僌儕儞傪妶惈壔偟偰嵶朎偺堏摦偺懀恑偵娭傢傞偙偲偑抦傜傟偰偄偨偺偱丄幉嶕偺惉挿墌悕偺怢挿偵丄R-Ras偑懀恑揑偵摥偒丄Plexin-B1偑偦偺妶惈傪梷惂偡傞偙偲偵傛傝丄斀敪嶌梡傪帵偡偺偱偼側偄偐偲悇掕偟偨丅偦偙偱丄Plexin-B1偺R-Ras偵懳偡傞GAP妶惈傪挷傋偨寢壥丄in
vivo媦傃in vitro偱R-Ras GAP妶惈傪帵偟丄偙偺妶惈敪尰偵Rnd1偺Plexin-B1傊偺寢崌偑昁恵偱偁傞偙偲偑傢偐偭偨丅傑偨丄懠偺掅暘巕検G抈敀幙丄H-Ras傗Rho僼傽儈儕乕G抈敀幙側偳偵懳偟偰偼慡偔GAP妶惈傪帵偝偢丄R-Ras偵摿堎揑側GAP
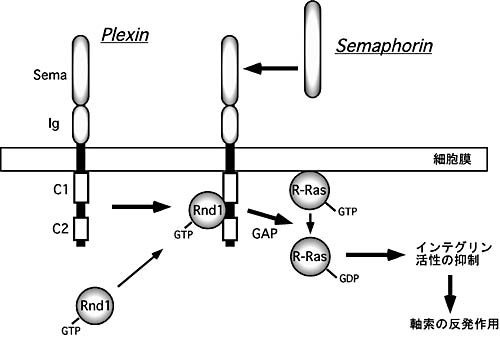
偱偁偭偨丅師偵丄Plexin-B1偺R-Ras GAP妶惈偑丄幉嶕偺斀敪嶌梡傪堷偒婲偙偡偺偐PC12嵶朎傗弶戙攟梴奀攏恄宱嵶朎偱挷傋偨丅Plexin-B1偺GAP妶惈傪寚懝偝偣偨曄堎懱傗Rnd1偺寢崌弌棃側偄曄堎懱偱偼Sema4D偵傛傞恄宱撍婲偺戅弅偑婲偒側偄偙偲丄傑偨丄忢帪妶惈宆偺R-Ras-QL傗siRNA偵傛傞撪嵼惈偺Rnd1僲僢僋僟僂儞偑Sema4D偵傛傞恄宱撍婲偺戅弅傗惉挿墌悕偺徚幐傪慾奞偟偨偙偲側偳偐傜丄恾侾偵帵偡傛偆偵Plexin-B1-Rnd1暋崌懱偵傛傞R-Ras
GAP妶惈偑Sema4D-Plexin-B1偵傛傞斀敪嶌梡偵昁梫偱偁傞偙偲偑傢偐偭偨丅Plexin-B1偺嵶朎撪椞堟偺R-Ras GAP僪儊僀儞偼懠偺Plexin偱傕傛偔曐懚偝傟偰偄傞偺偱丄Plexin-B偲嫟偵傛偔尋媶偝傟偰偄傞Sema3A-Plexin-A偺僔僌僫儖偵偍偗傞R-Ras
GAP偺娭梌傪挷傋丄弶戙攟梴奀攏恄宱嵶朎偵偍偄偰Sema3A偵傛傞惉挿墌悕偺徚幐傕R-Ras偺妶惈梷惂偑昁恵偱偁傞偙偲偑傢偐傝丄R-Ras
GAP偑Plexin僼傽儈儕乕偺晛曊揑側忣曬揱払宱楬偱偁傞壜擻惈偑悇嶡偝傟偨丅
|
恾侾丏Sema-Plexin偵傛傞幉嶕斀敪嶌梡偺嵶朎撪忣曬揱払宱楬
Plexin偺嵶朎撪椞堟丄C1偲C2偼R-Ras偵懳偡傞GAP偱偁傝丄Rnd1偑寢崌偟偨暋崌懱偱R-Ras妶惈傪梷惂偟丄恄宱幉嶕偺斀敪嶌梡傪敪婗偡傞丅Sema丗Sema條僪儊僀儞丄Ig丗僀儉僲僌儘僽儕儞條僪儊僀儞 |
丂俁丄壽戣丒揥朷
丂R-Ras偺嵶朎婡擻偲偦偺暘巕婡峔偼偁傑傝傛偔暘偐偭偰偄側偄偑丄嵶朎枌偺怢揥傗嵶朎偺慜恑側偳偵廳梫側栶妱傪壥偨偟偰偄傞偙偲偼娫堘偄側偄偲巚傢傟傞丅Plexin偼恄宱嵶朎傪娷傔偨條乆側嵶朎偺塣摦傪惂屼偟偰偄傞偙偲偑帵偝傟偰偄傞偺偱丄幉嶕偺斀敪嶌梡偵壛偊丄恄宱嵶朎偺堏摦偺惂屼傕R-Ras
GAP妶惈傪夘偟偰峴傢傟偰偄傞偙偲偑廫暘偵梊憐偝傟傞丅偲偙傠偱嵟弶偵弎傋偨傛偆偵丄Plexin偺嵶朎撪椞堟偺C1偲C2偺R-Ras GAP僪儊僀儞偼庬傪挻偊偰傛偔曐懚偝傟偰偄傞偺偱丄R-Ras
GAP妶惈偑Plexin偺斀敪嶌梡敪尰偺庡梫側忣曬揱払婡峔偱偁傞偲梊憐弌棃傞丅偟偐偟丄C1偲C2偵嫴傑傟偨Rnd1偺寢崌椞堟偼Plexin僼傽儈儕乕偺拞偱偁傑傝曐懚偝傟偰偍傜偢丄偝傜偵丄僔儑僂僕儑僂僶僄傗慄拵偵偼Rnd1偵懳墳偡傞G抈敀幙偑懚嵼偟側偄偺偱丄Rnd1偵傛傞Plexin偺R-Ras
GAP妶惈偺挷愡婡峔偼歁擕椶側偳偵尷嬊偝傟偨傕偺偺壜擻惈偑崅偄丅偦偺傛偆側Plexin偱偼丄懠偺Rho僼傽儈儕乕G抈敀偐丄偁傞偄偼慡偔暿偺妶惈挷愡婡峔偑娭傢偭偰偄傞偺偐傕偟傟側偄丅尰嵼傑偱偵丄幉嶕僈僀僟儞僗暘巕偺庴梕懱傪娷傔丄憹怋場巕傗暘壔場巕側偳偺懡偔偺庴梕懱偺嵶朎撪忣曬揱払婡峔偑柧傜偐偵偝傟偰偒偨偑丄掅暘巕検G抈敀幙偺GAP傪捈愙僐乕僪偟偰偄傞庴梕懱偼Plexin偑弶傔偰偺働乕僗偱偁傞丅崱屻丄條乆側幉嶕僈僀僟儞僗暘巕偺庴梕懱偺婡擻偑暘巕儗儀儖偱柧傜偐偵偝傟傞偙偲偵傛傝丄暋嶨側恄宱夞楬宍惉偺巇慻傒偑師戞偵柧傜偐偵側偭偰偔傞偩傠偆丅
暥專
1乯 Pasterkamp R J, Kolodkin A L. Semaphorin junction: making tracks
toward neural connectivity. Curr Op Neurobiol 13: 79-89, 2003
2乯 Swiercz J, Kuner R, Behrens J, Offermanns S. Plexin-B1 directly interacts
with PDZ-RhoGEF/LARG to regulate RhoA and growth cone morphology. Neuron
35: 51-63, 2002
3) Negishi M, Katoh H. Rho family GTPases as key regulators for neuronal
network formation. J Biochem 132: 157-166, 2002
4) Oinuma I, Katoh H, Harada A, Negishi M. Direct interaction of Rnd1
with Plexin-B1 regulates PDZ-RhoGEF-mediated Rho activation by Plexin-B1
and induces cell contraction in COS-7 cells. J Biol Chem 278: 25671-25677,
2003
5) Ishikawa Y, Katoh H, Negishi M. A role of Rnd1 GTPase in dendritic
spine formation in hippocampal neurons. J Neurosci 23: 11065-11072,
2003
6) Scheffzek K, Ahmadian M R, Wittinghofer A. GTPase-activating proteins:
helping hands to complemet an active site. Trends Biochem Sci 23: 257-262,
1998
|