
 |
GATA-1堚揱巕偺敪尰惂屼椞堟偺尋媶
|
|
|
|
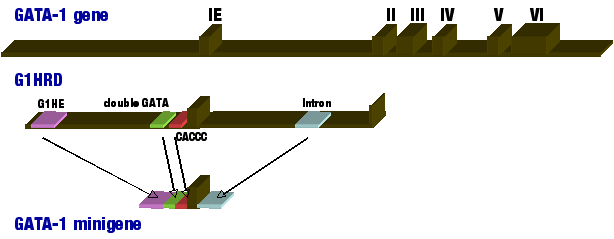
丂揮幨場巕GATA-1偼丆GATA攝楍乮WGATAR乯傪擣幆偟偰DNA偵寢崌偡傞揮幨挷愡場巕GATA僼傽儈儕乕乮GATA-1偐傜GATA-6偺俇偮偺場巕偵傛傝峔惉偝傟傞乯偺傂偲偮偱丆寣媴宯偺嵶朎偱偼丆愒夎媴丆嫄妀媴丆岲巁媴丆旍枮嵶朎側偳偵敪尰偟偰偄傞丏摿偵GATA-1偑僌儘價儞堚揱巕傗僿儉崌惉峺慺側偳偺愒寣媴暘壔偵廳梫側堚揱巕傗丆懡偔偺嫄妀媴娭楢堚揱巕偺敪尰傪摑崌揑偵惂屼偟偰偄傞偙偲偐傜丆巹偨偪偼丆尞場巕GATA-1偺敪尰惂屼婡峔傪専摙偡傞偙偲偑丆屄懱偱偺愒寣媴嫄妀媴憿寣僔僗僥儉傪棟夝偡傞偨傔偵昁恵偲峫偊偰偄傞丏 丂GATA-1堚揱巕偵偼東栿偝傟側偄戞堦僄僋僜儞偲丆幚嵺偵抈敀傪僐乕僪偡傞僄僋僜儞偺丆寁俇偮偺僄僋僜儞偐傜峔惉偝傟偰偄傞丏巹偨偪偼丆帋尡娗偺拞偱偼嵞尰偱偒側偄丆恀偺敪尰惂屼僔僗僥儉傪夝愅偡傞偨傔偵丆GATA-1偺敪尰惂屼椞堟偵兝僈儔僋僩僔僟乕僛乮LacZ乯丆椢怓寀岝抈敀乮GFP乯,敪岝峺慺乮Luc乯側偳偺儗億乕僞乕堚揱巕傪峔抸偟丆僩儔儞僗僕僃僯僢僋儅僂僗傗僩儔儞僗僕僃僯僢僋僼傿僔儏傪嶌惢偟偰夝愅偟偰偄傞丏偙傟傑偱偵丆IE偺忋棳-3.9 kbp傛傝戞俀僄僋僜儞傑偱偺椞堟偑偁傟偽丆寣媴宯偵墬偗傞惗棟揑側GATA-1堚揱巕偺敪尰傪嵞尰偡傞偺偵廫暘偱偁傞偙偲傪尒偄偩偟偨乮恾1乯丏巹偨偪偼丆偙偺椞堟傪G1HRD : GATA-1 hematopoietic regulatory domain偲屇傃丆GATA-1偺寣媴暘壔偵偍偗傞GATA-1偺婡擻婡擻傪屄懱偱専徹偡傞偨傔偺僣乕儖偲偟偰梡偄偰偄傞乮GATA-1偲寣媴暘壔偺崁傪嶲徠乯丏傑偨丆偙偺椞堟撪偺徻嵶側夝愅偵傛偭偰丆戀帣宆愒寣媴偱偺GATA-1堚揱巕敪尰偵偼丆忋棳傛傝G1HE椞堟丆double GATA椞堟丆CACCC椞堟偑廳梫偱偁傝丆惉懱宆愒寣媴偱偺GATA-1偺敪尰偵偼丆偙傟傜俁偮偺椞堟偵壛偊偰丆僀儞僩儘儞椞堟偺惂屼椞堟偑昁恵偱偁傞偙偲丆偙傟傜係偮偺僔僗椞堟傪僞儞僨儉偵楢寢偟偨栺侾kb偺峔抸乮GATA-1 minigene乯偱丆屄懱偱偺GATA-1偺敪尰傪嵞尰偱偒傞偙偲傪尒偄偩偟偨乮恾1乯丏嫽枴怺偄偙偲偵丆GATA-1堚揱巕偺庡梫側惂屼椞堟偺懡偔偑丆GATA box傪娷傓椞堟偱偁偭偨丏偙偺偙偲偼丆GATA-1傪娷傓GATA揮幨場巕孮偑GATA-1堚揱巕偺敪尰傪惂屼偟偰偄傞偙偲傪帵偟偰偄傞丏幚嵺丆儗億乕僞乕僩儔儞僗僕僃僯僢僋僼傿僢僔儏傪梡偄偨夝愅偐傜丆帺屓擇検懱傪宍惉偟偨GATA-1偑丆GATA-1帺恎偺堚揱巕敪尰椞堟偵寢崌偟丆偦偺敪尰傪惓偵惂屼偟偰偄傞偙偲傪柧傜偐偵偟偰偄傞丏 |
|
 |
||
|
恾侾
|
||
|
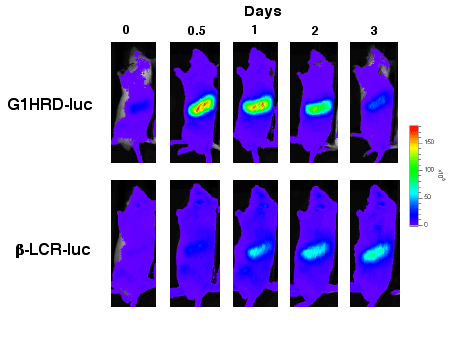
丂偲偙傠偱丆摦暔屄懱偐傜憿寣慻怐偲傝偩偟偰夝愅偡傞廬棃偺夝愅曽朄偱偼丆屄懱枅偺宱帪夝愅偑峴偊側偐偭偨丏偦偺寚揰傪夞旔偡傞偨傔偵偼丆儖僔僼僃儔乕僛敪岝峺慺傪梡偄偨僩儔儞僗僕僃僯僢僋儅僂僗偼桳岠側僣乕儖偵側傞丏巹偨偪偼丆敪岝峺慺傪僐乕僪偡傞堚揱巕傪GATA-1偺敪尰惂屼椞堟乮G1HRD乯偍傛傃兝僌儘價儞敪尰惂屼椞堟乮兝-LCR乯偵楢寢偟丆偦傟偧傟僩儔儞僗僕僃僯僢僋儅僂僗傪嶌惉偟偨丏昻寣桿摫傗掅巁慺忦審壓側偳偺僗僩儗僗憿寣帪偵偼丆傑偢寣拞偺僄儕僗儘億僄僠儞擹搙偑忋徃偟丆偦傟傪僩儕僈乕偲偟偰GATA-1堚揱巕偑敪尰偟丆傗傗抶傟偰僌儘價儞堚揱巕偺敪尰偑忋徃偡傞偙偲傪尒偄偩偟偨丏尰嵼偼丆偙傟傜愒寣媴憿寣偵廳梫側場巕偺敪尰傪儕傾儖僞僀儉偱夝愅偡傞偙偲偱丆憿寣偺峆忢惈堐帩婡峔偺偝傜側傞夝柧偵庢傝慻傫偱偄傞丏 |
||
 |
||
| 恾俀丏GATA-1偺敪尰惂屼椞堟乮G1HRD乯偲b僌儘價儞偺敪尰惂屼椞堟乮b-LCR乯偵儖僔僼僃儔乕僛敪岝婎幙傪僐乕僪偡傞堚揱巕傪楢寢偟丆僩儔儞僗僕僃僯僢僋儅僂僗屄懱撪偱偺偦傟偧傟偺堚揱巕敪尰傪専摙偟偨丏掕忢忬懺偱偼丆偄偢傟偺敪尰傕専弌姶搙埲壓偱偁偭偨偑丆昻寣偐傜偺夞暅帪偵偼丆傑偢GATA-1堚揱巕乮忋抜乯偑敪尰偟丆抶傟偰b僌儘價儞堚揱巕乮壓抜乯偑敪尰偡傞丏 |