
 |
憿寣宯偱摥偔丆揮幨場巕GATA-2
|
|
|
|
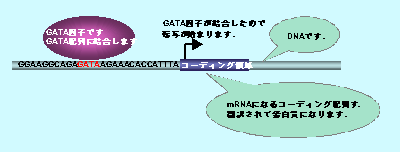
堚揱巕偺敪尰挷愡偲偼丆揮幨場巕偲偼 堚揱巕偼丆mRNA傪嶌傞僐乕僨傿儞僌椞堟偲丆偦偺堚揱巕偺敪尰挷愡偵摥偔僲儞僐乕僨傿儞僌椞堟偐傜側偭偰偄傑偡偑丆僲儞僐乕僨傿儞僌椞堟偺乽GATA乿攝楍傪擣幆偟偰寢崌偟丆偦偺廃埻偵偁傞僐乕僨傿儞僌椞堟偐傜mRNA傪嶌傜偣傞乮偁傞偄偼嶌傞偺傪巭傔偝偣傞乯巜椷傪弌偡偺偑丆揮幨場巕偱偁傞GATA場巕偱偡丏偙偺傛偆偵丆偝傑偞傑側揮幨場巕偑丆偦傟偧傟DNA偺摿暿側攝楍傪擣幆丆寢崌偟丆廃埻偺mRNA偺敪尰傪妶惈壔偟偨傝丆梷惂偟偨傝偟偰偄傑偡丏 |
|
 |
||
| 揮幨場巕GATA-2
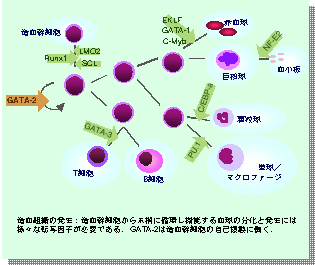
僸僩傗儅僂僗偱偼俇庬椶偺GATA場巕偑偁傝傑偡丏寣媴宯偱摥偔GATA場巕偼丆GATA-1, 2, 3偱偡丏GATA-2偼丆憿寣宯慻怐偺懠丆恄宱傗斿擜婍宯偺敪惗偵摥偄偰偄傑偡偑丆傢偨偟偨偪偼丆憿寣宯偱偺GATA亅俀偺栶妱偵偮偄偰丆尋媶傪恑傔偰偒傑偟偨丏 憿寣慻怐偺敪惗偼丆尩枾偵僾儘僌儔儉偝傟偰偄傑偡丏恾偼丆堦偮偺憿寣姴嵶朎偐傜丆慜嬱嵶朎偲屇偽傟傞拞娫揑側嵶朎傪宱偰丆懱撪偱堎側偭偨婡擻傪扴偆丆宍懺揑偵傕堎側偭偨條乆側寣塼嵶朎偑暘壔偟偰偄偔條巕傪帵偟偰偄傑偡丏偙偺暘壔偑恑傓偨傔偵偼丆奺僗僥僢僾偵偍偄偰懡悢偺揮幨場巕偑僾儘僌儔儉捠傝偵敪尰偟丆懠偺抈敀幙傪師乆偲寁夋捠傝偵敪尰偝偣偰偄偔偙偲偑昁梫側偺偱偡丏 GATA-2偺堚揱巕偑攋夡偝傟偨僲僢僋傾僂僩儅僂僗偱偼丆戀帣婜憗婜偵巰傫偱偟傑偄傑偡偑丆堦搙敪惗偟偨憿寣姴嵶朎偑丆堐帩偝傟憹怋偡傞抜奒偵堎忢偑偁傞偙偲偑傢偐偭偰偄傑偡丏俀杮偁傞堚揱巕偺曅曽偩偗偑攋夡偝傟偨応崌丆惗偒墑傃傞掱搙偵忈奞偼寉偄偺偱偡偑丆傗偼傝惉懱偵側偭偰傕丆崪悜憿寣偼丆惓忢儅僂僗偲斾妑偡傞偲丆忈奞偝傟偰偄傑偡丏
丒丒丒偙偺傛偆側栤戣堄幆偺傕偲偵丆傢偨偟偨偪偼尋媶傪恑傔偰偄傑偡丏 |
||
 |
||
|
|
||
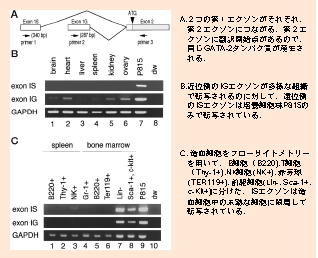
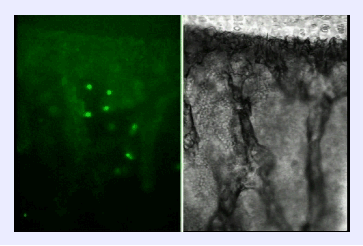
| GATA-2偺堚揱巕峔憿丂乕寣媴宯偵摿堎揑偵敪尰偡傞戞侾僄僋僜儞傪敪尒 崱偼慡僎僲儉偺忣曬偑僨乕僞儀乕僗偵搊榐偝傟偰偄傑偡偑丆埲慜偼丆偦偆偄偭偨傕偺偼偁傝傑偣傫偱偟偨丏GATA-2傪尋媶偡傞偨傔偵偼丄GATA-2偺堚揱巕峔憿傪柧傜偐偵偡傞偙偲偑昁梫偱偟偨丏 儅僂僗GATA-2堚揱巕偺峔憿偼丆俀偮偺戞侾僄僋僜儞偑偁傝丆偦傟偑偳偪傜傕戞俀僄僋僜儞偵偮側偑傝傑偡丏東栿奐巒揰偼戞俀僄僋僜儞偵偁傞偺偱丆嶻惗偝傟傞抈敀幙偼摨偠側偺偱偡偑丆揮幨偺巒傑傞偟偔傒偑丆俀偮偺僄僋僜儞偱偼堎側偭偰偄傑偡丏僸僩偵偍偄偰傕丆傗偼傝戞侾僄僋僜儞偑俀偮偁傞偙偲傕柧傜偐偵偟傑偟偨丏 嫽枴怺偄偙偲偵丆嬤埵懁偺戞侾僄僋僜儞乮General first exon; IG exon)偼條乆側慻怐偱揮幨偝傟偰偄傞偺偵斾傋丆墦埵懁偵偁傞戞侾僄僋僜儞 (Specific first exon; IS exon) 偼憿寣姴嵶朎偵嬤偄丆Lin-乮暘壔偟偨嵶朎偵弌偰偄傞儅乕僇乕偑堿惈乯暘夋傗丆Sca-1+, c-Kit+暘夋乮傗偼傝枹弉側嵶朎傪懡偔娷傓乯偺傒偱敪尰偑擣傔傜傟傑偟偨丏 Minegishi et al. http://www.jbc.org/cgi/content/full/273/6/3625 |
||
 |
||
| GATA-2偑憿寣敪惗晹埵偱偁傞Aorta-Gonad-Mesonephros (AGM)偱敪尰偟偰偄傞
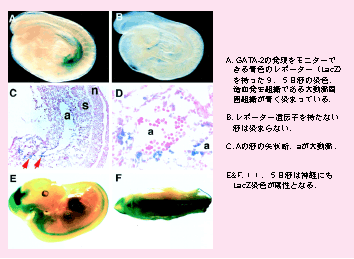
懕偄偰丆GATA-2偺弶婜憿寣敪惗偱偺敪尰傪柧傜偐偵偟傑偟偨丏In situ hybridization朄傗丆IS僄僋僜儞傪棙梡偟偨LacZ儗億乕僞乕傪梡偄偨僩儔儞僗僕僃僯僢僋儅僂僗傪嶌傞偙偲偱丆偦傟傜偑柧傜偐偵側傝傑偟偨丏IS僄僋僜儞傪娷傓GATA-2 堚揱巕惂屼椞堟偵丆LacZ愼怓偵傛偭偰惵偔敪怓偡傞儗億乕僞乕堚揱巕乮LacZ堚揱巕乯傪偮側偓丆僩儔儞僗僕僃僯僢僋儅僂僗傪嶌惢偟傑偟偨丏LacZ堚揱巕傪敪尰偝偣傞偟偔傒偼丆GATA-2傪敪尰偝偣傞偟偔傒傪巊偭偰偄傞偨傔丆LacZ愼怓偱惵偔敪怓偡傞慻怐偱偼丆GATA-2偑敪尰偟偰偄傞偲峫偊傜傟傑偡丏恾偺A偱惵偄晹暘偼丆戝摦柆傗惗怋婍丆彨棃恡憻偵側傞拞恡乮Aorta-Gonad-Mesonephros 丟AGM乯偲偄偭偨慻怐偑敪惗偡傞晹暘偱偁傝丆屄懱敪惗偺夁掱偱丆嵟弶偵懱撪偱寣塼偑嶌傜傟傞晹暘偲峫偊傜傟偰偄傑偡丏椢怓偵岝傞寀岝抈敀幙乮GFP乯傪儗億乕僞乕堚揱巕偲偟偰巊梡偡傞偲丆GATA-2傪敪尰偟偰岝傞嵶朎偑摼傜傟傑偡丏嵶朎昞柺偺摿挜傪夝愅偡傞僼儘乕僒僀僩儊僩儕乕偲偄偆曽朄傪梡偄傞偲丆GFP傪敪尰偡傞嵶朎偼丆枹弉側寣塼嵶朎偺惈幙傪帩偮偙偲偑傢偐傝傑偟偨丏 Minegishi et al. http://www.bloodjournal.org/cgi/content/full/93/12/4196 |
||
 |
||
| 崪悜拞偺憿寣姴嵶朎傪壜帇壔偱偒傑偟偨両
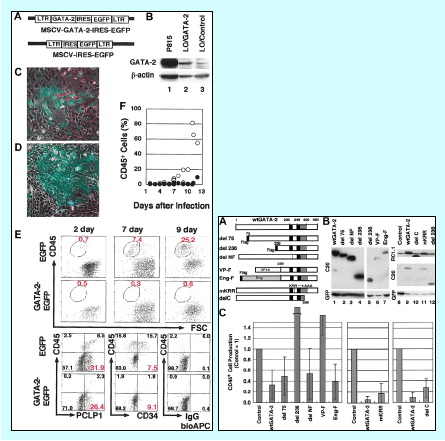
僩儔儞僗僕僃僯僢僋儅僂僗傛傝傕丆傛傝惓妋偵GATA-2偺敪尰傪棟夝偡傞偨傔丆GATA-2堚揱巕偺応強偵GFP傪擖傟傞偙偲偱丆GATA-2敪尰傪儌僯僞乕偱偒傞儅僂僗傪嶌惢偟傑偟偨丏乮Suzuki, N et al. www.pnas.org/cgi/doi/10.1073/pnas.0508928103) 姴嵶朎偼旝彫娐嫬偲偺憡屳嶌梡偵傛傝堐帩偝傟偰偍傝丆憿寣姴嵶朎傕崪悜偺側偐偱僯僢僔僃偲尵傢傟傞旝彫娐嫬偵埶懚偟偰懚嵼偡傞偲尵傢傟偰偄傑偡丏傢偨偟偨偪偼丆GATA-2偺憿寣宯偵摿堎惈偺崅偄IS僄僋僜儞偵椢怓寀岝抈敀幙乮GFP乯傪僲僢僋僀儞偟偨丆僲僢僋僀儞儅僂僗傪梡偄偰丆崪悜拞偺憿寣姴嵶朎偺摦夋嶣塭偵悽奅偱弶傔偰惉岟偟傑偟偨丏 恾偼丆偦偺儅僂僗偺崪悜偱偡丏塃懁偑偲摨偠椞堟傪寀岝尠旝嬀偱嶣塭偟偨傕偺偑嵍懁偺恾偱偡丅椢怓偵嫮偔岝傞嵶朎偑丆憿寣姴嵶朎偱偡丏憿寣姴嵶朎偼崪慻怐偵増偆條偵偟偰丆偟偐傕堦偮堦偮偑偍屳偄偵棧傟偰懚嵼偟傑偡丏嫽枴怺偄偙偲偵丆廃埻偺寣塼嵶朎偑妶敪偵摦偄偰偄傞偺偵丆憿寣姴嵶朎偼偠偭偲摦偒傑偣傫丏 偙偺寀岝傪栚報偵丆僼儘乕僒僀僩儊僩儕乕偺庤朄傪梡偄偰丆憿寣姴嵶朎偩偗傪庢傝弌偡偙偲偑壜擻偱偡丏偙傟傜偺嵶朎傪傕偭偲徻偟偔挷傋傟偽丆堦偮堦偮偺憿寣姴嵶朎偑丆偄偮暘楐偟偰偄傞偺偐丠偦偺帪丆崱傑偱偲摨偠乽憿寣姴嵶朎乿偵側傞偺偐丠偁傞偄偼憹怋丒暘壔偟丆幚嵺偵懱拞偱栶偵棫偮乽寣塼嵶朎乿傊堢偭偰偄偔偺偐丠偦偆偄偭偨嵶朎偼偳傫側摿挜傪傕偭偰偄傞偐丠偙偺儅僂僗傪梡偄偰丆憿寣姴嵶朎偺怴偟偄巔偑晜偐傃忋偑偭偰偔傞偱偟傚偆丏 |
||
 |
||
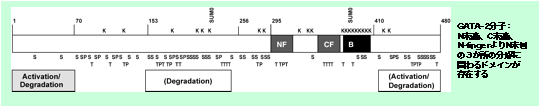
| GATA-2偺暘巕夝朥亅侾丂揮幨梷惂壔僪儊僀儞 師偼丆GATA-2偺拞偵栚傪岦偗傑偡丏 抈敀幙偺拞偱丆晹暘乮僪儊僀儞乯偛偲偵丆堎側傞婡擻傪暘扴偟偰偄傑偡丏GATA-2偼揮幨場巕偱偡偺偱丆DNA偵寢崌偡傞晹暘偲丆揮幨傪妶惈乛梷惂壔偡傞晹暘偑偁傞偼偢偱偡偑丆崱傑偱揮幨妶惈壔偺徻偟偄巇慻傒偼傢偐偭偰偄傑偣傫偱偟偨丏
|
||
 |
||
| GATA-2偺暘巕夝朥亅2丂儐價僉僠儞壔
GATA-2偑婡擻偡傞嵶朎偱偼丆GATA-2傪嶌傞偙偲乮敪尰乯丆GATA-2傪夡偡偙偲乮暘夝乯偺椉曽傪捠偠偰丆GATA-2偺ON/OFF僗僀僢僠偑挷愡偝傟偰偄傞偲峫偊傜傟傑偡丏偦偙偱丆GATA-2偑夡偝傟傞帪偵偼偳偺傛偆側僔僌僫儖偑偳偙偵擖傞偺偐丆傪挷傋傑偟偨丏 仺壓恾偵偼丄N僼傿儞僈乕乮NF乯偲C僼傿儞僈乕乮CF乯傪帩偮GATA2抈敀幙傪帵偟偰偄傑偡丅GATA-2偵偼N枛抂丄C枛抂丄N僼傿儞僈乕偺N懁偲丄俁偮偺暘夝偵娭傢傞僪儊僀儞偑偁傞偙偲偑柧傜偐偵側傝傑偟偨丏DNA偵寢崌偡傞N僼傿儞僈乕偲C僼傿儞僈乕偼懠偺GATA場巕偲斾妑揑帡偰偄傑偑丆暘夝偵娭傢傞僪儊僀儞偼偙傟傜偼懠偺GATA場巕偲憡摨惈偑掅偄晹暘偱偁傝丆GATA1偲GATA-2偱偺暘夝婡峔偺堘偄偑偁傞偲峫偊傜傟傑偡丏N枛抂懁偺暘夝僪儊僀儞偼丆揮幨妶惈壔婡擻傕帩偭偰偄傑偡丏儐價僉僠儞壔偲揮幨妶惈壔擻偲偺娭楢偼懠偺揮幨場巕偱傕巜揈偝傟傞偲偙傠偱偁傝丆偙偺傛偆側暘夝偲寢傃偮偔抈敀幙偺廋忺偑丆摨帪偵壓棳堚揱巕偺揮幨妶惈壔傪傕偨傜偡暘巕婡峔偺夝柧偼嫽枴怺偄壽戣偱偡丏 Minegishi et al. Genes to Cells, 2005 |
||
 |
||
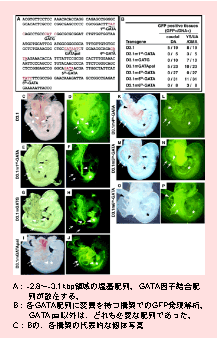
| 揮幨場巕GATA-2帺恎偺揮幨挷愡婡峔
GATA-2帺恎偺揮幨挷愡偺偟偔傒傪扵傞偙偲偼丄GATA-2偺敪尰傪巜椷偟偰偄傞揮幨場巕傪抦傞偙偲偵偮側偑傝傑偡丅GATA-2偺敪尰傪奐巒偝偣傞僔僌僫儖偼丠偦傟偼憿寣慻怐偺敪惗傗丄堦搙偱偒偨憿寣姴嵶朎偺帺屓暋惢丄憹怋偵偳偺傛偆偵娭傢偭偰偄傞偺偱偟傚偆丠 GATA-2偺揮幨偼丄旕忢偵峀偄揮幨挷愡椞堟偵傛偭偰挷愡偝傟偰偄傑偡丅嫟摨尋媶幰偺Engel嫵庼偺尋媶幒偱偼丄戝挵嬠恖岥愼怓懱乮BAC) 傪梡偄偰丄GATA-2堚揱巕偺忋棳傗壓棳偺悢侾侽侽僉儘墫婎懳 (kbp) 偺斖埻偺夝愅傪恑傔偰偄傑偡丅憿寣偵娭傢傞揮幨挷愡椞堟偼丄250 kbp偺BAC傪巊偭偰柧傜偐偵偝傟傑偟偨丅 傢偨偟偨偪偼丄GATA-2偺IS僄僋僜儞忋棳3.1 kbp偺斖埻偵丄憿寣宯偱摥偔揮幨挷愡椞堟偑偁傞偙偲傪柧傜偐偵偟傑偟偨丅3. 1bbp忋棳堟傪丄GFP偺揮幨妶惈壔椞堟偲偟偰梡偄偨堚揱巕傪梡偄偰僩儔儞僗僕僃僯僢僋儅僂僗傪嶌惉偡傞偲丄戀帣偺憿寣慻怐 (AGM側偳乯偑GFP偺敪尰偵傛傝椢怓偵岝傝傑偡丅偦偟偰丄僸僩偲儅僂僗偲偺娫偱墫婎攝楍憡摨惈偑旕忢偵崅偄丄-2.8乣3.1 kbp 偺斖埻偺夝愅傪恑傔偰偄傑偡丅 塃恾A偼丄-2.8乣3.1kbp偺椞堟偺墫婎攝楍丄B偼D3.1峔抸偵彮偟偢偮曄堎傪擖傟偨攝楍偺僩儔儞僗僕僃僯僢僋儅僂僗夝愅偺寢壥丄C偼B偱帵偟偨奺乆偺峔抸偵偮偄偰丄戙昞揑側屄懱偺幨恀傪帵偟偰偄傑偡丅傑偢丄A偵帵偡捠傝丄-2.8乣-3.1kbp椞堟偵偼丄GATA場巕寢崌攝楍偑俇偐強偁傝傑偡丅偦偟偰丄偦偺奺乆偵曄堎傪擖傟偨峔抸偱僩儔儞僗僕僃僯僢僋儅僂僗傪嶌惢偡傞偲丄傕偟傕丄曄堎傪擖傟偨GATA攝楍偑廳梫偱偁傟偽丄GFP偺岝傝曽偑曄傢偭偰偒傑偡丅D3.1峔抸偼丄戀帣婜偺憿寣慻怐偱偁傞戝摦柆偲棏墿擷丄娞憻尨婍偑傂偐傝傑偡偑丄B傪傒傞偲丄憿寣慻怐偱偁傞戝摦柆偱GFP偑岝傞偨傔偵偼丄GATApal埲奜偺攝楍偼偳傟傕昁梫偱偁傝丄GATA-2偺敪尰偵偲偭偰偼丄偄偢傟偐偺GATA場巕偑偙偺椞堟偵寢崌偡傞偙偲偑昁梫側偙偲偑傢偐傝傑偟偨丅 偦偺懠丄 -2.8乣3.1kbp椞堟拞偵偼偦偺懠偺揮幨場巕偑寢崌偡傞岓曗偲側傞傛偆側攝楍偑懡悢懚嵼偟丄偦偺堦偮堦偮偵偮偄偰丄夝愅傪恑傔偰偄傑偡丅 Kobayashi-Osaki et al. Mol Cell Biol 2005 http://mcb.asm.org/cgi/content/full/25/16/7005?view=long&pmid=16055713 |
||
 |
||
| GATA-2偵偮偄偰偺暥愑偼彫椦丂杚偱偡丏偛幙栤偼maki_k@tara.tsukuba.ac.jp傑偱偍婑偣偔偩偝偄丏 |