| A02班 公開シンポジウム報告書
柚崎通介(慶應大学・医学部・生理学)
A02班では「脳の発生生理機能の研究」をテーマに、特に視覚野、感覚野、小脳をモデルとして、神経栄養因子、グルタミン酸受容体、Caチャネルなどの分子に焦点をあて、津本、中村、渡辺各計画班員に加えて、広く公募班員が募集して活発な研究が行われてきた。まず、津本班長から、研究成果のまとめが報告され、引き続き狩野班員および鍋倉班員の研究成果が報告された。紙数の関係ですべては触れられず、また一部誤解があるかも知れないが、どうかご容赦いただきたい。
1)まとめ(津本班長)
本班では、主に、発達期における、(1)シナプスの選択的強化と除去過程、(2)受容体・神経伝達物質・トランスポータの変換過程、についての研究が進められた。
(1)では、津本班長らは、大脳視覚野における眼優位性カラム形成過程における脳由来栄養因子(BDNF)の作用機序について、BDNFがシナプス前部から後部へ、順行性に放出されて取り込まれる過程を緑色蛍光蛋白(GFP)との融合蛋白を用いたイメージングにより明らかにした(Science,
291, 2419-2423, 2001)。従来考えられていた標的細胞由来の逆行性シグナルに対して、順行性シグナルとしてもBDNFが作用することを発見した画期的な成果である。さらに、BDNFノックアウトマウス由来と、GFP発現マウス由来の神経細胞を混合培養することにより、このような順行性シグナルが、抑制性神経の発達に特異的に作用することを明らかにした(J
Neurosci 23, 6123-6131, 2003)。
哺乳類脳における速い神経伝達は、AMPA型グルタミン酸受容体により担われているが、幼若期の脳においては、NMDA型グルタミン酸受容体しか存在しない「サイレント」シナプスが多く存在し、神経活動に依存してAMPA受容体を含むシナプスに変化することが知られている。中村班員らは、感覚野(バレル)の脳切片を用いて、BDNFノックアウトマウスではこの過程が障害され、かつBDNFの投与により回復することから、サイレントシナプスの成熟過程がBDNFにより制御されていることを初めて発見した(PNAS
100, 13069-13074, 2003)。
小脳プルキンエ細胞は、幼若時には多数の登上線維により支配され、次第に1:1の支配様式に変化する。P/Q型Caチャネル欠損マウスにおいては、この成熟過程が遷延することが、渡辺班員と狩野班員らにより示され、小脳登上線維の選択的強化・除去過程におけるCaチャネルの役割が初めて明らかとなった(J
Neurosci 24, 1734-43, 2004)。(後述する狩野班の紹介の項も参照のこと)
(2)では、GABA-A受容体は、成熟脳では抑制性神経伝達を担うが、幼若脳では、細胞内Clイオン濃度が高いために、興奮性に働くことがこれまでに知られていた。福田班員らは、グラミジシンパッチ法と単一細胞RT-PCR法を用いることにより、その原因が、幼若皮質神経細胞ではClトランスポータNKCC1の発現量が多いためであることを明らかにした(J
Physiol 557:829-41, 2004)。発達に伴う抑制性神経伝達の変化については、後述のように鍋倉班員らが、伝達物質そのものがGABAからグリシンに変化することを発見した(Nature
Neurosci 7, 17-23, 2004)。
|
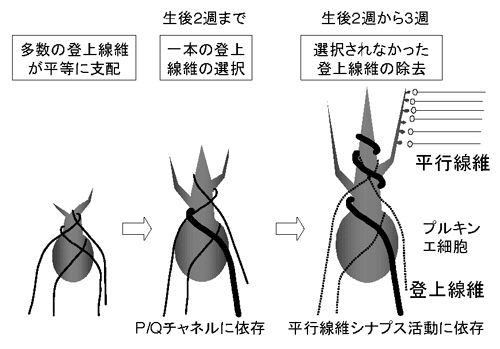
図1. 登上線維によるプルキンエ細胞の1:1支配様式の生後発達過程 |
2)「発達期小脳におけるシナプスの刈り込みと機能成熟(狩野班員)
幼若時の小脳では、数本の登上線維がプルキンエ細胞を支配するが、生後発達に伴い、げっ歯類では生後24日令(P24)くらいまでに、約90%のプルキンエ細胞において1対1の支配様式が確立する。この系は、発達期におけるシナプス刈り込みと機能成熟過程を検討するためには非常に有用である。狩野班員らは、(1)どのようにして一本の登上線維が選択され、(2)どのような過程で残りの登上線維シナプスが除去されるか、についてこれまでの研究成果を総括した。
P3での小脳では、複数の登上線維がほぼ同じくらいの伝達効率で、一個のプルキンエ細胞を支配する。その後P12前後までに、このうちの一本の登上線維のシナプス伝達効率が選択的に強化され、シナプス間の反応のばらつきが増大し、次第に小さい反応を示すシナプスが選択的に除去される。狩野班員らは、個々の登上線維シナプスにおけるグルタミン酸放出量を、低親和性のグルタミン酸受容体拮抗剤PDAを用いることにより推定し、小さい反応を示すシナプスでは、登上線維からのグルタミン酸放出量が小さいことを発見した(Neuron
38, 785-96, 2003)。即ち、シナプスの位置や放出様式の変化などにより、一回の興奮で放出されるシナプス小胞の数が少なくなった登上線維が、やがて選択的に除去されることを示唆する。
一方、「先端脳」以前の研究により、狩野班員らは、これまでにNMDA型グルタミン酸受容体の拮抗剤MK-801を、P15〜P16の間に小脳皮質へ局所的投与すると、余剰登上線維の除去過程が特異的に阻害されることを発見した(J
Neurosci 20:4954-61, 2000)。また、同様の登上線維の多重支配現象は、平行線維-プルキンエ細胞シナプスにおける代謝型グルタミン酸受容体mGluR1やその下流の信号分子であるPLC(PNAS
95, 15724-9, 1998)やGq(PNAS 94, 14089-94, 1997)の遺伝子欠損マウスでも存在することから、登上線維-プルキンエ細胞シナプスの除去は、小脳皮質における平行線維-プルキンエ細胞シナプスの活動に依存する異シナプス性機構により遂行されると考えられている。今回、狩野班員らと渡辺班員らは、P/Q型Caチャネル遺伝子欠損マウスにおいては、P12に至っても、登上線維のシナプス伝達効率の選択的強化が観察されないことを発見した。すなわち、登上線維の選択的除去過程には2つの相があり、生後2〜3週において選択されなかった弱い登上線維を除去する異シナプス性の過程に先行して、生後2週までに一本の登上線維のシナプス伝達を選択的に強化する過程が存在し、この過程にはP/Q型Caチャネルの活性が必要であることを示唆する(J
Neurosci 24, 1734-43, 2004)。
3)「抑制性神経伝達物質の発達スイッチング」(鍋倉班員)
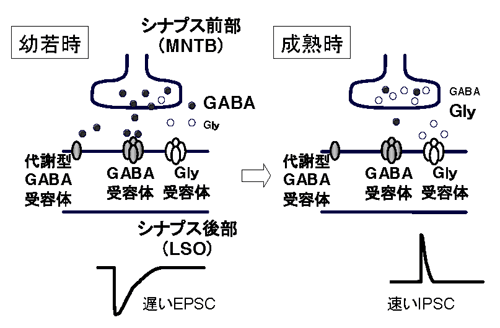
外側上オリーブ核(LSO)は、同側の蝸牛核より興奮性の聴覚入力を受け、反対側の蝸牛核より台形体内側核(MNTB)を介して抑制性の入力を受けることにより、左右の聴覚情報の僅かな差異を統合して、音がどの方向から来ているか(音源定位)に関与している。これまでに、LSOへのMNTB入力の伝達物質は、発達段階によりGABAからグリシンに変化することが報告されていた。しかし、この現象が果たしてGABA作動性神経細胞が除去されて、グリシン作動性神経細胞に置換されるのか、あるいは個々の神経細胞において伝達物質そのものが置換されるのかについては不明であった。鍋倉班員らは、神経前終末部が付着した状態のままで、LSO神経細胞を急性単離し、微小抑制性シナプス後電流mIPSCを測定することにより、個々の神経前終末部において、伝達物質がGABAからグリシンに置換されることを明確に示した(Nature
Neurosci 7, 17-23, 2004)。また電顕や光顕レベルにおいてもこの所見は裏付けられた。
これまでに発達段階に伴う抑制性神経回路のリモデリングの研究は、興奮性のそれに比べて遅れていた。しかも、神経伝達物質そのものが発達段階に伴って変化するという現象はまったく新しい原理であり、GABAとグリシンが混在する他の脳部位における抑制性神経回路にも当てはまる可能性があり、非常に興味深い。この現象の生理的意義としては、前述のようにGABAやグリシンは幼若期の神経細胞においては、興奮性伝達物質として作用するので、より遅い反応を示すGABAは、幼若期の神経細胞においては、可塑性の誘導に必要なCaレベルの上昇には有利である可能性がある。また、GABAは、グリシンとは異なり、GABA-B受容体を介した代謝型反応を活性化する可能性もあり、鍋島班員らは実際に、幼若期LSOにおいて、GABA-Bを介した反応が見られることを報告した。一方、蝸牛を破壊すると、成熟後にもGABA→グリシン変換が起きないことから、これらの現象は神経活動に依存したものであることも示唆された。
|
図2. 生後発達に伴うGABAからGlyへの伝達物質の変化 |
4)感想
私は、これまで長年米国にいた為に、「班会議」型式の研究費システムは今年が全く初めてである。正直なところ、狭い日本において、通常の学会や各種シンポジウムに加えて、班会議のような型式での班員間の交流や情報交換の機会の必要性があるのかどうか、懐疑的な気持ちがあった。同時に、このような型式が日本で定着するには、何かしらの利点や理由がったはずであり、批判をする前に、もう少しこのシステムについて知る必要があると思っている。したがって、現時点での私の感想は瑣末的なことがらにとどめる。
公開シンポジウムや夏のワークショップは、新たなことを勉強できて有用であった。他の班員の進捗具合を知るのは刺激になってよかったが、今回の班会議のような5分発表・質疑なしという口頭発表型式は、私には辛かった。むしろポスター型式のみで、もう少しゆっくりと時間を費やしたかった。口頭発表でやるならば、せめて発表・質疑を入れて20〜30分は欲しい。また、夏のワークショップのように、折角泊りがけで、班会議をしたのならば、数日間宿泊し、例えばGordonやKeystoneシンポジウムなどで行われているような、研究者同士のインフォーマルな交流機会を増やすことが、新たな有機的な研究の発展を生む土台になると思う。今年の夏のような懇親会では、あまりにも巨大で、薄い交流しかできなかった。とはいえ、今回の班会議も夏のワークショップも、企画する側の先生方にとっては大変なご負担と尽力であると思う。最後に、心から感謝の意を表し、今後の「統合脳」の発展を祈りたい。
|