| B03班公開シンポジウム報告書
モデル脳による記憶・学習・思考の研究
林 初男(九州工業大学 大学院生命体工学研究科)
平成12年度から始まった特定領域研究「先端脳」は最終年を向かえ、昨年12月21日に砂防会館で、5年間の総まとめとしての公開シンポジウムが行われた。5年間の総まとめであるので、7つすべての班による発表が行われた。どの班の研究成果も優劣つけがたいすばらしいものであり、先端的かつ濃密な研究発表を聞かせていただき、普通の研究会では感じられないような質の高さに感銘を受けた。
さて、B03班の研究項目は、「モデル脳による記憶・学習・思考の研究」である。脳のニューロン集団の活動がダイナミカルな現象であることを踏まえると、脳ではダイナミカルな情報表現がなされ、それに基づいた学習や記憶がなされていると考えるのは自然である。B03班の研究目的は、そのような立場から脳理論を確立するために、学習・記憶のダイナミクスモデルを構築し、脳の学習・記憶・思考の理論的解明を行うことであった。
| |
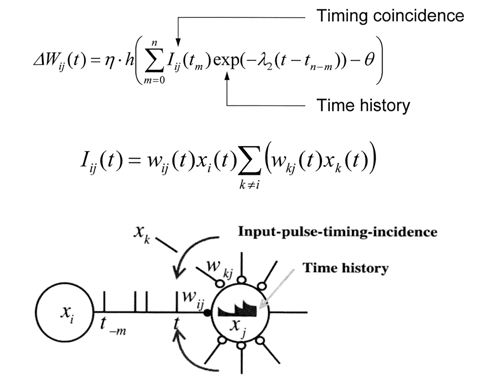
図1. 時空間学習則。この学習則はスパイクタイミング依存性と時間的履歴効果の両方を考慮した多細胞間の学習則。学習の結果、時空間情報を反映したシナプス荷重の空間分布が形成される。 |
時空間情報がニューロン活動の時空間ダイナミクスで表現されているとすれば、学習の結果としても、時空間情報が神経回路網のシナプス荷重空間の上に表現されなければいけない。そこで、塚田らは、図1に示すような、時空間情報をシナプス荷重空間に埋め込むことのできる時空間学習則を提唱した1)。この時空間学習則はスパイクタイミング依存性と時間的履歴効果の両方を考慮した多細胞間の学習則である。細胞間の発火タイミングと時間的加重特性に基づいてシナプス結合強度が変化し、時空間情報を反映したシナプス荷重の空間分布が形成される。しかし、時空間学習則は仮説であるので、その妥当性の実験による検証や時空間情報の学習記憶機構としての有効性などを明らかにする必要があった。
塚田らは、オプティカルレコーディング法やマルチユニット解析法を用いて、可塑的に変化するシナプス荷重の空間分布等を調べ、時空間学習則の実験による検証を行った。まず、海馬スライスを用い、シャファー側枝が走っているCA1放線状層を刺激した。1ヵ所を刺激してもLTPは生じないが、2ヵ所同時に刺激するとLTPが生じ、刺激のタイミングを20
ms程度ずらすと、刺激の順序には関係なくLTDが生じた。これは、入力−入力間のタイミングに依存したシナプスの可塑的変化が生じていることを示している。次に、CA1放線状層の1ヵ所に時系列刺激(マルコフ連鎖)を加え、マルコフ連鎖の相関に依存したLTPが誘導されることを示した。これは、LTP誘導に入力信号の時間的履歴効果があることを示している。さらに、CA1錐体細胞の樹状突起基部と末端部にシナプスを作っているシャファー側枝をいろいろなタイミングで刺激すると、樹状突起基部ではメキシカンハット型のSTDP関数が得られ、末端部では非対称なSTDP関数が得られた。GABAA受容体阻害剤であるビキュキュリンを投与すると、CA1錐体細胞の樹状突起基部で見られるSTDP関数はメキシカンハット型から非対称なSTDP関数に変わるので、メキシカンハット型STDP関数は抑制性介在ニューロンによるものであり、抑制性介在ニューロンによる抑制が少ない樹状突起末端部では非対称なSTDP関数となることが明らかになった。このようなネットワークの構造に依存する入力−出力タイミング依存性の違いはシナプス荷重の空間分布形成に大きな影響を与えることになる。塚田らは、さらに、シナプス後細胞の発火に依存しない入力−入力タイミング依存性LTPが生じることも明らかにした。以上の実験結果は、時空間学習則の妥当性を検証したものである。塚田らは、最後に、時空間学習則による学習は、Hebb則による学習に比べ、時系列信号パターンを区別する能力が高いことをモデルシミュレーションで明らかにし、時空間学習則が時空間情報の学習記憶機構として有効であることを示した。
時空間学習則に関わるその他の研究成果としては、以下に示すものが報告された。津田は、海馬CA3領野のカオス活動が海馬CA1領野の安定な神経回路に入力されたとき、CA1ニューロンの応答から再構成したアトラクタに、カントール集合と呼ばれる階層構造が生じることを明らかにした2)。この階層構造に情報を書き込みかつ保持させることができるので、カオスダイナミクスの中に情報を埋め込めることが明らかになった。林らは、海馬CA3神経回路網モデルのリカレント結合にスパイクタイミングに依存するシナプス可塑性を持たせ、CA3神経回路網の局所領域にθバースト信号を加えた。その結果、加えた位置から放射方向に興奮性結合が強化され、ニューロン活動の放射状伝搬がθ周波数で繰り返すことを見出した3)。さらに、CA3神経回路網の複数の異なる局所領域に時間をずらしてθバースト信号を加えると、各局所領域から波紋状に広がる興奮波のダイナミカルな空間パターンが生じ、これらのCA3ニューロンの発火とCA1へ貫通路を通じて入力されるθバースト信号とのタイミングに依存してシャファー側枝シナプス結合強度が可塑的に変化した。その結果、入力された信号の時間的順序関係をコードしたシナプス荷重の空間分布が生じた。このようにシナプス荷重の空間分布を形成する学習は、塚田が提唱している時空間学習則に通じるものである。
以上の一連の研究成果と密接に関連しながら、合原らは、数理モデル解析により、集団発火率コーディングや集団時間コーディングといったパラダイムで脳の学習記憶理論を展開した。そのようなニューロン集団によるコーディング機構を考える場合、ニューロン活動の同期や非同期といったダイナミカルな現象とその機能的役割の解明が重要になる。
| |
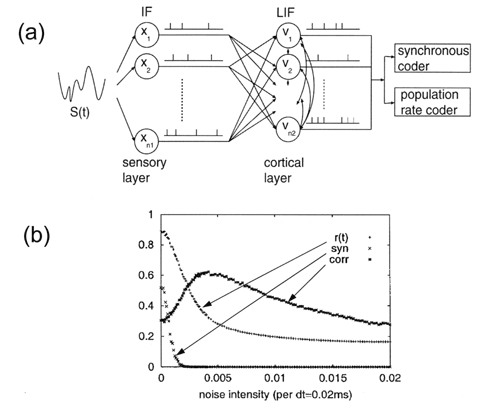
図2. 同期コーディングと集団発火率コーディング。
(a) フィードフォワードネットワークモデル。IF:Integrate-and-fire neuron。LIF:Leaky
integrate-and-fire neuron。第2層のニューロンには雑音が加えられている。(b) オーダーパラメータおよび相関係数の雑音強度依存性。syn
(×)は1個のニューロンの発火数に対する同期発火数の割合。
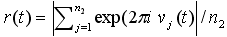
ただしn2は第2層のニューロン数、νj
(t) はj 番目のニューロンの膜電位。corr (*)は、入力信号と第2層の集団発火率との相関係数。 |
合原らは、図2(a)に示すような2層のネットワークを用い、ニューロン集団が集団発火率コーディングと同期コーディングの両方のコーディング機構を持ちうることを示し、図2(b)に示すように、集団発火率コーディングと同期コーディングのどちらの機構が有効になるかは、第2層のニューロンに加わる雑音の強さに依存することを明らかにした4)。十分弱い雑音を加えると発火が同期する傾向になり、同期間隔に信号をコードできる。それに対し、ある程度強い雑音を加えると発火は非同期になり、集団発火率の時間変化として入力信号の概形をコードできる。もちろん、雑音だけが集団発火率コーディングと同期コーディングを切り替える方法ではなく、フィードフォワードやフィードバック結合の局所性、フィードバックの強さ、積分発火型ニューロンのリークレート、あるいはニューロン特性のばらつきなども2つのコーディング機構を切り替える方法として使える。このように、ニューロン集団は集団発火率コーダーとしても同期コーダーとしてもはたらくので、多層フィードフォワードネットワークとして、メキシカンハット型結合を有するSynfire
Chainを用いると、同期発火の伝搬だけではなく、集団発火率の伝搬を起こすこともでき、集団発火率の伝搬から同期発火の伝搬へと変化させることもできる。さらに、複数の入力信号がフィードフォワードネットワークに加えられたときのコーディングについても調べた。STDPルールに従うシナプスを通じて、複数のポアソンランダムパルス列をネットワークに加えると、STDPによるシナプス競合によってアクティブニューロンのクラスターが形成され、それらのクラスターが集団発火コーダーあるいは同期コーダーとしてはたらくことを明らかにした。
次に、2つのコーディング機構がはたらいているとき、生理学的エルゴード性が成り立っているかどうかが問題になる。単一ニューロンの繰り返し入力に対する応答の試行平均とニューロン集団の単一入力に対する応答の集団平均とが等価になるとき生理学的エルゴード性が成り立っているという。平均膜電位がしきい値に近ければ発火間隔の分布はシャープになり、そのピークは入力信号間隔のヒストグラムのピークにロックされる。これを試行同期(trial
synchrony)と呼んでいる。合原らは、エルゴード性が成り立つかどうかはこの試行同期性と集団同期性の兼ね合いで決まることを明らかにした5)。すなわち、試行同期と集団同期が同時に生じたときエルゴード的であり、試行非同期と集団非同期が同時に起きたときは、発火率に関してエルゴード的である。雑音の効果として調べると、雑音が十分小さいとき試行同期と集団同期が実現され、発火のダイナミクスはエルゴード的である。雑音強度がある程度大きくなると、試行同期がくずれるので、エルゴード的ではなくなる。雑音が十分強くなると、集団同期も崩れるが、集団発火率が入力信号の波形をかなり忠実に再現するので、エルゴード性が復活する。
合原らは、最後に、興奮性GABAニューロンの動的記憶システムに及ぼす影響を明らかにしているが、本報告の紙面が限られているので、省略させていただきたい。
先にも述べたように、「先端脳」5年間の総まとめとして行われた公開シンポジウムで、各班から報告された研究成果は目を見張るすばらしいものばかりであったと思う。しかし、5年間の総まとめであるこの公開シンポジウムで、唯一つ物足りなく感じた点がある。「先端脳」は一つの特定領域として組織された研究プロジェクトである。7つの班の研究成果がどのように有機的に結びつき、先端脳全体としては何が確立できたのか、あるいはこれからの脳研究の方向性として何が示せたのかなど、ある程度時間をかけた総括講演がほしかったところである。そうすることにより、「先端脳」の成果がこれからの脳研究に大きな指針を与えることができ、また、これから始まる「統合脳」に対しても、より高い山に挑むための礎になっていることを明確にできたのではないかと思う。とはいうものの、充実した発表内容や研究最前線にふさわしい質疑応答を聞いて、各班の5年間の研究成果のすばらしさを実感したことは偽りのないところである。
B03班の5年間のまとめの報告では、発表時間の関係で、公募研究として参加された方々の研究についてはあまり詳しい報告はなされなかった。しかし、多くの優れた研究成果をあげられたことを最後に申し添えたい。また、「統合脳」においても、脳研究のいろいろな側面で、脳のダイナミクス理論がさらに発展することを期待したい。
参考文献
1) Tsukada M, A computational model of learning and memory. Int. Cong.
Series 1269 (2004) 11-20.
2) Tsuda I and Kuroda S, Cantor coding in the hippocampus. Jpn. J. Ind.
Appl. Math. 18 (2001) 249-258.
3) Yoshida M and Hayashi H, Regulation of spontaneous rhythmic activity
and organization of pacemakers as memory traces by spike-timing-dependent
synaptic plasticity in a hippocampal model. Phys. Rev. E 69 (2004) 011910:
1-15.
4) Masuda N and Aihara K, Bridging rate coding and temporal spike coding
by effect of noise. Phys. Rev. Lett. 88 (2002) 248101.
5) Masuda N and Aihara K, Ergodicity of spike trains: When does trial
averaging make sense? Neural Comp. 15 (2003) 1341-1372.
|