| B01班 「記憶・学習・思考の分子生物学的研究」
平野丈夫(京都大学・大学院理学研究科)
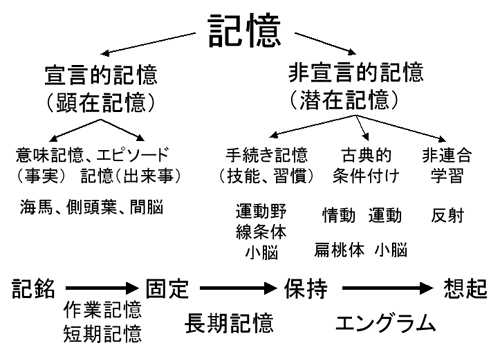
B01班の発表では、班長の三品昌美教授が研究班活動のオーバービューといくつかの成果の概略を紹介し、続いて神戸大学の饗場篤教授が最新の研究成果を報告した。B01班は、図に示したような様々な記憶の諸過程においてシナプス機能分子がどのようにはたらいているかを探求することを通じて、記憶・学習の分子機構を理解することをめざしてきた。
B01班ではシナプス・細胞内での機能分子の動態を探る研究やさまざまなミュータントマウスを用いてシナプス機能分子の動物個体内でのはたらきを調べる研究等がなされ、また両者を結びつけることも試みられてきた。研究手法も多岐にわたり、マウスを用いて時期および脳部位特異的に遺伝子ノックアウトを行う手法、特定神経細胞の選択的ノックアウト、シナプス部位での機能分子動態のイメージング、二光子蛍光顕微鏡でケージト伝達物質を利用して個々のシナプスを選択的に刺激する手法等、最新技術の開発や採用がなされてきた。三品氏は、自身の研究室で行ってきた標準的なマウス系統C57BL/6を用いて時期および脳部位選択的に遺伝子をノックアウトする手法等を用いたミュータントマウスについての研究、福島県立医科大学の小林和人氏グループの選択的神経細胞ノックアウトマウスを用いた大脳基底核神経回路に関する機能解析、高知大学の椛秀人氏グループの匂い記憶に関する分子・細胞生理学的研究、生理学研究所の松崎政紀氏グループの単一スパインレベルでの長期増強発現機構についての研究(News
Letter 5巻2号に紹介記事)、私の研究室で行ってきた小脳抑制性シナプス可塑性制御の分子機構の解析等、を紹介した。この発表により、B01班の多彩で活発な研究活動の一端が明らかにされたように思う。
饗場氏は、メタボトロピックグルタミン酸受容体mGluR1ノックアウトマウスにおいて、プルキンエ細胞特異的にmGluR1サブタイプを発現させて、機能回復を解析する一連の実験結果を報告した。メタボトロピックグルタミン酸受容体のなかで、mGluR1はmGluR5とともにグループ1を形成し、三量体G蛋白質Gqと共役しフォスフォリパーゼC活性制御を介し、プロテインキナーゼC活性や細胞内カルシウムイオン濃度を調節している。mGluR1にはスプライシングの違いにより、長い細胞内C末端を有するmGluR1aと短い細胞内C末端しかもたないmGluR1b,
cのサブタイプが存在することが知られている。饗庭氏はmGluR1ノックアウトマウスにおいて、L7プロモーターを用いて小脳プルキンエ細胞選択的にmGluR1aを発現させると、ノックアウトマウスで見られたプルキンエ細胞への登上線維の多重支配、平行線維・プルキンエ細胞間シナプスでの長期抑圧欠損、運動失調が認められなくなることを示した。この結果は、上記mGluR1ノックアウトマウス表現型がプルキンエ細胞でのmGluR1の働きに依存していたことを示している。
今回、饗場氏はmGluR1ノックアウトマウスにおいてmGluR1bをプルキンエ細胞特異的に発現させる実験を行った。mGluR1bは、Gqと共役可能でありフォスフォリパーゼCも活性化できるが、mGluR1aの細胞内C末端に存在するHomer/Veslとの結合に必要なモチーフやシナプス後部の足場蛋白質群との結合に関与するPDZ結合モチーフはもたず、それらのシナプス後部分子との結合能が劣っていると推定される。今回、mGluR1bレスキューマウスを作成してその表現型の解析を行った。まず、mGluR1b蛋白質の免疫沈降物にHomer/Veslが含まれないことを確認した。次に、mGluR1bレスキューマウスでローターロッドテストを行い、運動制御能力が野生型マウスと同等にまで回復したことを示した。しかしながら、このレスキューマウスでは登上線維のプルキンエ細胞多重支配および瞬膜反射条件付けのdelay課題という学習試験については回復が認められなかった。長期抑圧については現在検討中ということであったが、学習の回復が認められないという結果は、長期抑圧が回復してない可能性を考えさせるものである。またレスキューマウスのプルキンエ細胞ではmGluR1アゴニスト投与により引き起こされる細胞内カルシウムイオン濃度上昇の持続時間が短いという結果も示した。これらの結果は、シナプス後部において足場蛋白質等との結合に関与すると考えられるmGluR1aの長い細胞内C末端は、プルキンエ細胞において運動協調に必要なシグナル伝達に必須でないが、登上線維の選択的除去、瞬膜反射の条件付け、細胞内カルシウム濃度の持続性上昇に必要であることを示唆している。
今回の結果は興味深く、いろいろな問題を提起しているように思う。私の第一の疑問は、mGluR1bはどのようにして運動協調を回復させたのであろうかという問題である。このレスキューマウスで、瞬膜反射の条件付けが回復しないということは運動学習能力が阻害されていることを示唆する。また登上線維のプルキンエ細胞多重支配も維持されているし、細胞内カルシウム応答は持続しない。Gqとの共役が回復したことにより、プルキンエ細胞のいかなる細胞機能が回復したのであろうか、それによって生体内でのプルキンエ細胞活動はどのように変化してそれが運動制御にどのように寄与したのであろうか。mGluR・Gqシグナル経路が野生型とは異なる形でもとにかく回復することが重要だったのであろうか。また、登上線維の多重支配が維持されても運動協調に問題がないとすると、野性型マウスで起こる登上線維の選択的除去の意味は何なのであろうか。学習効率への寄与であろうか。さらに、このミュータントマウスにおいて運動学習能力に欠失があるとすると、それが運動協調に影響しないというのはどういうことなのであろうか。運動協調能力に影響しない運動学習能力は動物にとって意味のあるものなのであろうか。それとも今回行われたローターロッドテストの条件では、運動制御能力測定の精度に限界があるということなのであろうか。より感度のよい運動制御能力テストでは差が検出できるのであろうか。また、運動学習〔長期抑圧?〕に必須なmGluR1のはたらきは何なのであろうか。細胞内カルシウムイオン上昇またはプロテインキナーゼCの活性化が持続されることなのであろうか。いろいろな点で考えさせられる興味深い報告であった。
私は発足当初から先端脳に参加してきたが、この5年間で様々なミュータントマウス作成を含む新たな実験手法の開発がなされ、幾多の共同研究が行われ着実に研究が進展して重要な成果が挙がったように思う。来年度から始まる統合脳においても、よい情報交換の場が提供され多様な研究者の育成がなされることにより、わが国の神経科学研究のさらなる活性化とレベルアップが行われることを祈っている。
|